










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Portuguesa de Ortopedia e Traumatologia
versão impressa ISSN 1646-2122versão On-line ISSN 1646-2939
Rev. Port. Ortop. Traum. vol.26 no.3 Lisboa set. 2018
ARTIGO DE REVISÃO
Mecanismos imunológicos na reabsorção espontânea de hérnias do disco intervertebral
Belmiro M. K. O. AlvesI; Jorge M. S. R. O. AlvesI
I. Faculdade de Medicina da Universidade do Porto.
RESUMO
Está comprovado que as hérnias do disco intervertebral podem sofrer um fenómeno de reabsorção espontânea e existe evidência na literatura que este processo ocorre por mecanismos imunológicos. O disco intervertebral é o maior órgão avascular do organismo, sendo considerado um dos locais imunologicamente privilegiados do ser humano. Perspetiva-se que a violação deste estado imunologicamente privilegiado ocorrida durante a herniação poderá expor o conteúdo do disco intervertebral ao sistema imune e assim ser desencadeada uma resposta imunológica a fim de proceder à sua reabsorção.
O objetivo desta “monografia” visa a revisão da literatura e a sistematização dos possíveis mecanismos pelos quais a reabsorção das hérnias do disco intervertebral acontece. A base de dados utilizada foi a PUBMED, onde foram selecionados artigos completos e revisões escritas em língua inglesa publicados entre 1953 e 2017. Os termos MeSH usados foram “intervertebral disc”, “intervertebral disc displacement”, “spontaneous resorption” “immune system”, isoladamente ou em combinação. A lista de referências dos artigos identificados foi também analisada na procura de mais artigos relevantes associados ao tema.
É consensualmente aceite que no processo de reabsorção das hérnias do disco intervertebral estão envolvidos fenómenos de neovascularização, quimiotaxia de células imunes e formação de um tecido de granulação, culminando num processo fagocitose do material herniado. Contudo, ainda existe controvérsia quanto ao tipo de imunidade implicada neste processo, pelo que quer a imunidade inata, quer a imunidade adaptativa poderão estar ambas implicadas.
Palavras chave: Hérnia de Disco Intervertebral, Disco Intervertebral, Sistema imunológico, Imunologia.
ABSTRACT
It has been proven that the intervertebral disc hernia may suffer a phenomenon of spontaneous resorption and there is evidence in literature that this is mediated by immunological mechanisms. The intervertebral disc is considered one of the human immunological privileged sites, being also the largest avascular organ in the human body. It is foreseen that the invasion of this immunologically privileged state that occurs during herniation may expose the IVD to the immune system hence triggering an immune response in order to promote its resorption.
The aim of this “monography” is to review literature and systematize the possible mechanisms by which the resorption of the intervertebral disc hernia occurs.
The database used was PUBMED where full text articles and English-written reviews published between 1953 and 2017 were preferred. The MeSH terms used were ‘intervertebral disc’, ‘intervertebral disc displacement’, ‘spontaneous resorption’ and ‘immune system’ either alone or in combination. The references of the articles were also considered when searching for the most relevant articles.
It is widely accepted that the resorption process of the herniated disc implies mechanisms such as neovascularization, chemotaxis of immune cells and synthesis of a granulated tissue culminating in the phagocytosis of the herniated material. However, there is controversy as to which immunity is implicated in this process thus both innate immunity and adaptive immunity may be involved.
Key words: Intervertebral Disc Displacement, Intervertebral Disc, Immune System, Immunology.
INTRODUÇÃO
A lombalgia e a dor ciática são uma das principais causas de incapacidade da população mundial, observando-se uma prevalência de 11,9%1. Aproximadamente 84% da população experiencia pelo menos um episódio de lombalgia ao longo da sua vida2. A hérnia do disco intervertebral (HDIV) é considerada como a principal causa de lombalgia e dor ciática3.
A evolução dos meios auxiliares de diagnóstico permitiu uma melhoria na avaliação e orientação de doentes com HDIV. Tornou-se evidente que em certos doentes estas regridem espontaneamente4-6. Numa meta-análise que incluía 11 estudos, Zhong et al. verificou que 67% das hérnias discais lombares são reabsorvidas espontaneamente7.
Normalmente a regressão espontânea associa-se ao alívio sintomático6. A capacidade de prever quais os doentes com maior probabilidade de regressão das HDIV vai permitir melhorar o aconselhamento do doente relativamente ao tratamento, evitando cirurgias desnecessárias, com morbilidade, riscos e custos conhecidos.
O disco intervertebral (DIV) é uma estrutura única, com capacidade de absorver choques e de resistir à deformação induzida pela carga. É composto por uma estrutura externa rígida, designada anel fibroso (AF), e uma estrutura interna gelatinosa, chamada núcleo pulposo (NP). O AF é composto principalmente por colagénio, enquanto o NP é constituído por proteoglicanos, maioritariamente agrecanos. Tem sido sugerido que o processo degenerativo se inicia no NP e depende da perda de proteoglicanos8. Geralmente, as HDIV ocorrem no contexto da degeneração do DIV9. Com as recorrentes agressões biomecânicas, aliadas à perda da homeostasia na composição do DIV, o AF acaba por romper, ocorrendo a falência da sua integridade e permitindo o extravasamento do NP10.
Ainda são desconhecidos os mecanismos concretos responsáveis pelo processo de reabsorção espontânea das HDIV. Várias hipóteses foram sugeridas, das quais realçamos a desidratação do tecido herniado, o repuxamento pelo ligamento longitudinal posterior (LLP) e o envolvimento de uma resposta imune. Independentemente destas hipóteses, foram encontrados vários fatores associados à maior probabilidade da ocorrência a regressão espontânea das HDIV.
MATERIAL E MÉTODOS
A base de dados utilizada foi a PUBMED, onde foram selecionados artigos completos e revisões escritas em língua inglesa publicados entre 1953 e 2017. Os termos usados foram “intervertebral disc”, “intervertebral disc displacement”, “spontaneous resorption” “immune system”, isoladamente ou em combinação. A lista de referências dos artigos identificados foi também analisada na procura de mais artigos relevantes associados ao tema.
DISCUSSÃO
1. Mecanismos de reabsorção hipotéticos
Uma das hipóteses propostas na literatura para a regressão espontânea das HDIV é a da desidratação, ocorrendo uma redução do tamanho do NP herniado após esse fenómeno11. Através de RMN, observa-se numa fase inicial que a intensidade de sinal do NP herniado aumenta em relação à do NP original, diminuindo de seguida com o tempo12. Esta perda de intensidade de sinal nas ponderações em T2 da RMN suporta a hipótese da desidratação12,13. Quando a intensidade de sinal é inicialmente elevada nas imagens em T2, a probabilidade da regressão do tamanho da hérnia é maior, comparativamente aos casos em que a intensidade de sinal inicialmente é baixa. Isto leva a inferir que a intensidade de sinal pode ser um bom preditor da redução espontânea do HDIV12,14. No entanto, a desidratação ao contribuir para uma redução do tamanho da HDIV, não explica os casos de desaparecimento completo do fragmento livre11.
Outra hipótese sugerida pressupõe a ocorrência de um fenómeno de retração mecânica do material herniado, retornando para o interior do DIV,4,15 possivelmente através de uma variação de tensão no LLP13 e também por um possível fenómeno de negativização da pressão do espaço intradiscal, levando ao repuxamento espontâneo do material herniado13,15. Contudo, como descreveremos mais à frente, a reabsorção espontânea das HDIV ocorre mais frequentemente quando estas se encontram sequestradas, estando neste caso totalmente separadas do próprio LLP16. Estes achados sugerem que a teoria do repuxamento do conteúdo herniado pelo LLP não se adequa na fundamentação de um mecanismo para a regressão espontânea de todas as HDIV.
Por último, a hipótese que relaciona o envolvimento do sistema imune no processo de reabsorção espontâneo das HDIV permanece a mais aceite pela comunidade científica. Conquanto, o próprio sistema imune é um vasto e complexo domínio gerador de múltiplas e distintas respostas, muitas vezes interligadas, que iremos relacionar com o seu possível envolvimento naquele processo. De um modo geral, permanece controvérsia na abordagem a este assunto pelos vários autores. A divergência de opiniões centra-se em duas hipóteses para o mecanismo imunomediado responsável pela reabsorção espontânea do DIV. Uma considera o envolvimento da imunidade inata, com a formação de uma resposta inflamatória direcionada ao NP. Outra foca a resposta secundária da imunidade adaptativa, com a formação de uma resposta autoimune direcionada ao NP.
2. DIV imunoprivilegiado
O DIV é a maior porção de tecido avascular do adulto, estando por esse motivo isolado do sistema imune do hospedeiro16,17. Ao ser considerado um dos locais imunoprivilegiados do ser humano, a existência de células imunológicas no seu interior é sinal de rotura da barreira imunológica. O ambiente imunologicamente privilegiado do DIV não se deve apenas a uma simples barreira física entre os tecidos do DIV e a circulação sanguínea, mas também à envolvência de uma série de mecanismos moleculares contribuidores para uma barreira fisiológica17,18.
O Fas Ligando (FasL) é expresso na superfície celular de linfócitos T citotóxicos (CTL), células Natural Killer (NK) e em locais imunologicamente privilegiados17. Atua causando a morte apóptica de células que expressam o seu recetor, por meio da ativação de uma cascata de caspases19. Foi comprovado que o FasL existe nas células do DIV, particularmente nas células do NP20. As células do NP saudáveis sobreregulam o FasL, induzindo a apoptose de células endoteliais vasculares, macrófagos e CTL que invadem o DIV17,18. Outro dado também relevante é que a expressão do FasL nas células do NP saudável é significativamente superior em relação ao NP degenerado17. Por outro lado, a expressão de FasL nas células do NP de HDIV contidas foi significativamente superior em relação às HDIV extrusas20.
Portanto, a expressão do FasL nas células do NP humano ao prevenir a neovascularização18 e a infiltração de células inflamatórias no DIV, através da indução da apoptose Fas-mediada17, mantem o estado avascular do NP e, consequentemente, a preservação do estado imunologicamente privilegiado, comprovando que existe uma barreira fisiológica suportada pela rede FasL-RecetorFas.
Outro fator que pode contribuir para um estado imunoprivilegiado do DIV é uma expressão de citocinas típicas de uma resposta do tipo Th2 pelas células do DIV. Esta evidência foi encontrada num estudo imunohistoquímico onde foi observado que as próprias células do DIV são positivas para os marcadores de linfócitos T e expressam citocinas típicas de uma resposta do tipo Th2 em HDIV contidas21.
Por outro lado, também foi descoberto que reações oxidativas em NP desnutridos podem levar à alteração fenotípica precoce das células residentes do NP, levando à expressão de propriedades fagocíticas. Nerlich et al. detetou células CD68+ em DIV degenerados sem hérnia associada, particularmente no NP e, em menor frequência, no AF. Morfologicamente, estas células CD68+ não diferem dos condrócitos do NP, assemelhando-se a células residentes transformadas com potencial fagocítico. É de destacar que foram observados aglomerados destas células particularmente em áreas de crescimento vascular22. Num outro estudo, Chen et al. observou células do NP fagocíticas com características intermédias entre macrófagos e células NP, possuindo uma maior quantidade de lisossomas que as células típicas do NP, assemelhando-se a histiócitos. Estes resultados sugerem que as células do NP, ao possuírem um potencial fagocítico capaz de eliminar células apoptóticas e porções de matriz extracelular danificada, podem contribuir para a manutenção do estado imunologicamente privilegiado do DIV23.
3. Grau da Herniação
Na tentativa de compreender os mecanismos responsáveis pela reabsorção espontânea das HDIV, diversos estudos procuraram estabelecer padrões e relacionar fatores que contribuem para este fenómeno.
Estudos recentes sugerem que o dano da plataforma cartilagínea predispõe o DIV à degeneração24, sendo mais severo nos DIV herniados comparativamente àqueles em que não ocorre herniação, sugerindo o possível envolvimento das plataformas cartilagíneas na patogenia das HDIV. Neste sentido, verificaram que a falência na HDIV ocorreria mais frequentemente devido a avulsão da plataforma cartilagínea do que por rotura do AF. O tamanho da porção avulsa não se correlacionou com o tamanho da herniação, podendo pequenas avulsões originar hérnias de grandes dimensões25. As lesões da plataforma cartilagínea são mais comuns em doentes com HDIV sintomáticas, estando associadas a uma maior percentagem de doentes com défices neurológicos e à diminuição das hipóteses de recuperação pelo tratamento conservador26.
As HDIV podem ser classificadas de acordo com o seu grau de severidade. Numa perspetiva crescente de gravidade, podem ser consideradas abaulamentos de disco, protusões (P), extrusões subligamentares (ES), extrusões transligamentares (ET) ou sequestros (S). De um modo geral, há uma maior tendência para a reabsorção espontânea com a evolução do grau da HDIV, sendo esta mais evidente nas ET e S5,16,27,28.
Fatores como o tamanho da hérnia, o grau de preservação do LLP, a extensão e a migração do seu conteúdo correlacionam-se significativamente com a diminuição do volume dos DIV herniados e a consequente reabsorção28. Dentro destes, o grau de preservação do LLP parece ser o que tem maior impacto como fator contributivo para a regressão espontânea da HDIV, distinguindo as ET e S das restantes27. Deste modo, tendo em conta a integridade do LLP, ao longo do presente trabalho consideraremos as P e ES como HDIV contidas (HDIVC), enquanto as ET e as S serão consideradas como HDIV extrusas (HDIVE), de modo a facilitar a compreensão das temáticas abordadas.
4. Neovascularização
A existência de neovascularização na periferia do NP herniado parece ser o maior determinante na reabsorção espontânea da HDIV29,30.
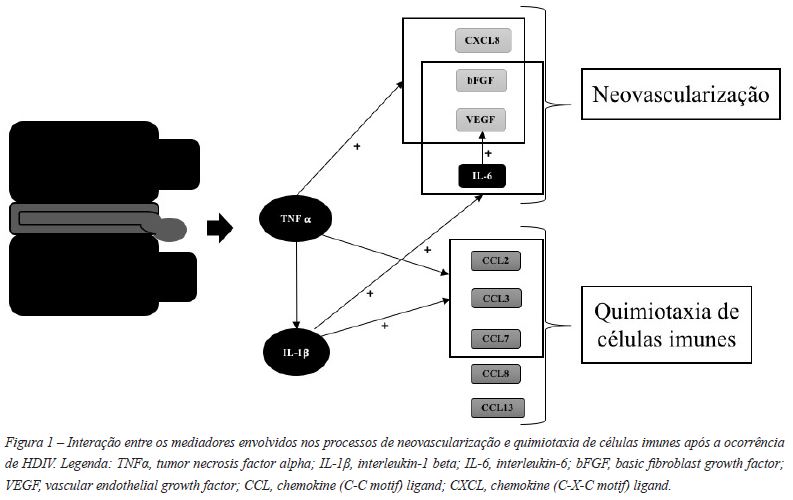
No sentido de compreender como ocorre o processo de neovascularização foram estudados os fatores angiogénicos responsáveis. Entre os quais damos um especial destaque ao bFGF e ao VEGF, deixando algumas notas acerca do CXCL8. De realçar que foram observadas associações entre as citocinas inflamatórias, nomeadamente o TNFα, a IL-1ß e a IL-6, e os fatores angiogénicos descritos30-33 (Figura 1) (Tabela 1).
O bFGF é expresso em condrócitos degenerados e em vasos sanguíneos formados nos tecidos de granulação32,34,35. O bFGF estimula a neovascularização e a quimiotaxia de macrófagos, linfócitos e fibroblastos32,36-38. Os níveis de bFGF relacionam-se diretamente com a neovascularização, sendo este um potente promotor da angiogénese32. Na verdade, promove o crescimento de vasos sanguíneos para o interior dos DIV lesados36, através da indução das células endoteliais capilares a invadir matrizes de colagénio, formando túbulos característicos de capilares sanguíneos38. A angiogénese dependente deste fator parece atuar em resposta à agressão ao DIV34. De facto, a expressão de bFGF associa-se à atividade proteolítica da matriz extracelular do DIV,34,35 pelo que é um dos mediadores no processo de reabsorção da HDIV37.
Relativamente ao VEGF foi demonstrado que ocorre um aumento da sua expressão em estádios precoces da HDIV39. Este é produzido pelas células do DIV e por macrófagos, enquanto as células endoteliais vasculares expressam os seus recetores40. A interação entre os macrófagos e os tecidos do DIV levam à indução do VEGF e à consequente estimulação da formação de mais vasos sanguíneos. Este fenómeno foi observado num estudo de cocultura de macrófagos e células do DIV onde a neovascularização foi mais abundante, em comparação à sua cultura isolada, o que demonstra a existência de uma sinergia entre elas para a sobreregulação daquele fator angiogénico40. Os macrófagos surgem no tecido de granulação decorrente do processo inflamatório verificando-se a existência de vasos sanguíneos a proliferar em seu redor. A angiogénese e a infiltração macrofágica aumentam com a proliferação de células produtoras de VEGF41, levando a concluir que o VEGF promove a quimiotaxia de monócitos40, originando um feedback positivo entre o número de macrófagos e a sua produção.
A expressão de células positivas para o VEGF e seus recetores no NP humano foi superior nos grupos de doentes com HDIV, sendo que nestes a expressão foi máxima em doentes com HDIV extrusa para o espaço epidural40.
A IL-6 é uma citocina proinflamatória que estimula o crescimento e a proliferação de vários tipos de células imunes durante as respostas imunológicas de defesa do organismo42 e contribui na neovascularização, nomeadamente através da sobreregulação do VEGF31, sendo produzida tanto pelo DIV como pelas células infiltradas no tecido de granulação43,44.
A CXCL8 produzida no DIV também parece ter um papel estimulador no crescimento de capilares sanguíneos para o interior dos tecidos da HDIV, facilitando a sua reabsorção, sendo que a sua produção é significativamente superior em doentes com HDIVE, principalmente nos S45.
Do ponto de vista anatómico, ao nascimento, o DIV é dotado de suprimento vascular, principalmente ao nível da plataforma cartilagínea e do AF. Contudo, esses vasos cedo sofrem regressão, deixando o DIV com um escasso ou nulo suprimento sanguíneo direto no adulto saudável46,47. A invasão do espaço do DIV por vasos sanguíneos pode facilitar a introdução de citocinas, fatores de crescimento, células e outras substâncias que influenciam a reabsorção tecidular e os subsequentes processos de reparação do DIV lesado36.
Porém, esta neovascularização não parece ser apenas produzida pela resposta inflamatória, mas também durante o processo degenerativo que precede em muitos casos a herniação9. Durante a degeneração do DIV ocorre um processo de neovascularização nas áreas circundantes do AF47,48, sendo este capaz de a induzir possivelmente devido ao seu conteúdo em substâncias solúveis indutoras da angiogénese. Tal não se constata na plataforma cartilagínea, o que pode ser o motivo pelo qual as HDIV com grande quantidade de plataforma cartilagínea não serem tão eficazmente absorvidas49. Esta evidência é suportada pelo estudo de Shan et al., que recorrendo à classificação de Modic50, onde são usadas as diferenças de intensidade de sinal das plataformas cartilagíneas na visualização por RMN, descreveu as diferenças na tendência para a reabsorção nas HDIV e os seus possíveis intervenientes. As hérnias dos grupos Modic Changes (MC) possuem mais cartilagem hialina, com menor conteúdo em NP e invasão vascular que os grupos não-MC. Nesse estudo foi também verificado uma maior abundância de capilares e macrófagos nas amostras de tecido herniado dos grupos não-MC. Os proteoglicanos, presentes em maior quantidade nos tecidos cartilaginosos, inibem a angiogénese, pelo que a sua perda encoraja a neovascularização e a tendência para a reabsorção do tecido herniado, demonstrada nos grupos não-MC48.
Na verdade, verifica-se a diminuição de proteoglicanos na maioria das HDIV, particularmente nas HDIVE, sendo esta mais proeminente nas porções adjacentes ao tecido de granulação51 e estando fortemente associada à neovascularização36,48.
Após a exposição do NP verifica-se uma neoformação de vasos sanguíneos, provavelmente devido à sua continuidade com o plexo venoso e tecido adiposo presentes no espaço epidural16,52. Mas o grau de neovascularização parece estar intimamente ligado com o tipo de HDIV53. As HDIVE são expostas com maior facilidade à circulação sanguínea e, consequentemente, a uma maior quantidade de células imunes, o que provavelmente leva à reabsorção por fagocitose dos tecidos herniados, existindo uma maior probabilidade de regressão espontânea da HDIV9,16,27,54,55. A infiltração no conteúdo herniado por vasos sanguíneos pode ocorrer através do LLP55, mas na verdade, quando a porção do NP da massa herniada perfura o LLP ocorre uma reação de neovascularização mais exuberante56. O conteúdo herniado é rodeado e infiltrado por vasos recém-formados a partir do tecido adiposo epidural, sendo posteriormente invadido por macrófagos e outras células do sistema imune27,37,55. Nesta perspetiva vários autores especulam que a neovascularização e a infiltração de células imunes na periferia de tecidos herniados, estejam implicadas nas diferenças observados entre os vários tipos de HDIV e no consequente processo de reabsorção5,9,16,27,37,51,54,55,57.
5. Quimiotaxia de células imunes
Na sequência do que foi descrito para a neovascularização e formação de tecido de granulação, Doita et al. verificou que ocorreria um aumento do número de células mononucleadas do sangue periférico ligadas à superfície de HDIVE, em comparação com HDIVC. Conclui que estas células mononucleadas, sobretudo monócitos, seriam essenciais no despoletar de uma resposta imunológica ao exercer um papel regulatório na produção de uma variedade de mediadores capazes de recrutar novos monócitos para as HDIV32,58. O DIV humano também parece ser dotado de propriedades quimiotáxicas, produzindo muitos destes fatores promotores do recrutamento de várias células imunes, chamados de quimiocinas45,59. Assim, o recrutamento de monócitos circulatórios para material herniado resulta da interação entre as quimiocinas produzidas pelo próprio IVD e a sua ligação a recetores da superfície dos monócitos. Os monócitos são assim ativados localmente, diferenciando-se em macrófagos19,60.
Portanto, numa primeira fase são as próprias células do NP a recrutar macrófagos através da produção de quimiocinas. É de destacar que a presença de macrófagos no tecido de granulação, na periferia do material herniado ou mesmo invadindo o NP e o AF, surge apenas após a expressão de quimiocinas sugerindo a necessidade destas no recrutamento de macrófagos59,61,62. Após infiltração nos fragmentos de disco herniado, os macrófagos vão também produzir quimiocinas com consequente aumento da sua população63. A produção conjunta dos fatores entre as células do NP e os macrófagos é mais relevante do que quando isolados, o que leva a crer que existe um feedback positivo entre ambos os elementos celulares, à semelhança do que já foi descrito para os fatores angiogénicos.
Estas quimiocinas parecem ser primariamente reguladas por citocinas proinflamatórias em resposta à agressão. A produção local de citocinas como a IL-1ß e o TNFα parece ser parte essencial do processo ao mediar indiretamente a regulação das quimiocinas envolvidas na quimiotaxia de células imunes63,64. Após a extrusão da HDIV, numa fase anterior à expressão de quimiocinas, o TNFα e a IL-1ß são produzidos de novo nas células do DIV, estimulando a expressão de quimiocinas43,62 (Figura 1).
Relativamente às quimiocinas propriamente ditas, tanto as células imunes infiltradas nos tecidos de granulação como as células do DIV são capazes de expressar CCL245,57,59,62,63,65, CCL357,66, CCL760,63 e CXCL843-45,63. Isoladamente as células do DIV expressam CCL863 e CCL1360 (Tabela 1).
Efetivamente, está comprovado o papel do CCL245,59,61,62,65, CCL357,66, CCL767, CCL867 e CCL1367 na capacidade de provocar o recrutamento e migração de células inflamatórias, pelo que as restantes quimiocinas poderão ter um papel regulatório intermediário no processo quimiotático. Há um aumento da expressão de CCL245,57, CCL357,66 e CXCL845 nas HDIV, pelo que estas parecem aumentar ainda mais com a severidade da HDIV57.
6. Resposta Imune
Os primeiros passos foram dados em 1952, quando Hirch et al. verificou a formação de um tecido de granulação altamente vascularizado após a rotura do AF com extrusão do seu conteúdo além do LLP. Observou a posterior constituição de uma cicatriz fibrosa no local lesado do AF46. Posteriormente em 1965, Bobechko et al. observou a ocorrência de uma reação imune do tipo corpo estranho à colocação de fragmentos de NP autógeno em locais vascularizados, com a produção de anticorpos68. Efetivamente, o perfil da ativação inflamatória e imunológica apresentada pelo HDIV parece variar desde a degeneração aos diversos graus de herniação69.
a. Imunidade Inata: perspetiva inflamatória
A resposta imune inata parece relacionar-se numap rimeira fase com uma reação inflamatória típica de corpo estranho em resposta à exposição do conteúdo do DIV. Como já foi referenciado previamente, o contacto entre o fragmento herniado do DIV e o espaço epidural parece ser o gatilho para despoletar a resposta imunológica através da infiltração de células inflamatórias e da neovascularização no local, com consequente formação de um tecido de granulação na periferia do tecido herniado54,61,65,70,71. Os vasos recém-formados têm um papel essencial na passagem de células inflamatórias infiltrativas. Os infiltrados celulares foram mais proeminentes no NP do que no AF54. Esta infiltração celular é composta por macrófagos54,57,62,65,72, neutrófilos62 e alguns linfócitos T54,57,62,65. Nestes estudos, raramente foram encontrados linfócitos B54,57,62,65.
Kagawachi et al. através de uma análise imunofenotípica de infiltrados inflamatórios de HDIV verificou que nenhum dos infiltrados continha linfócitos, monócitos ou células dendríticas, enquanto todos os infiltrados examinados continham macrófagos em abundância73.
A infiltração celular com formação do tecido de granulação é acompanhada de fibrose e relaciona-se inversamente com o tamanho dos DIV herniados62, existindo uma tendência na redução de tamanho com o tempo65.
A citotoxicidade das células imunes em relação às células do NP também foi alvo de estudo. Murai et al., cultivou células autólogas de NP juntamente com células imunes do mesmo indivíduo e verificou que a citotoxicidade para as células autólogas do NP foi proporcional à quantidade de células NK e macrófagos. Linfócitos T CD4+ e linfócitos T CD8+ não obtiveram efeitos citotóxicos nas células NP. Tendo em conta a citotoxicidade das células imunes, os alvos moleculares e a sua localização nas células NP permanecem indefinidos. De seguida, testaram a sobrevivência das células NP transplantadas em ratinhos imunocomprometidos e em ratinhos com o sistema imune preservado. Constataram que passados 3 meses um pequeno número células transplantadas sobreviveu no grupo imunocomprometido, enquanto nenhuma sobreviveu passadas 3 semanas no grupo com o sistema imune preservado. Não se verificou a migração de linfócitos T para as células NP, contudo observou-se a migração de células NK e macrófagos. Esta evidência demonstra que o NP é sensível a células imunes. Neste estudo os macrófagos não mostraram ser células residentes transformadas, mas células infiltrativas. Em conclusão, as células do NP provocam uma resposta imune inata, pelo que ao reconhecer o NP autólogo transplantado os macrófagos e as células NK parecem ter uma função imunológica precoce quando o NP é exposto ao sistema imune74.
Está demonstrado que há uma implicação da imunidade inata, nomeadamente através de uma resposta inflamatória no processo de reabsorção das HDIV. A maioria dos autores considera que esta reação inflamatória se deve à exposição do NP ao sistema imune, com uma consequente reação tipo corpo-estranho. Contudo alguns autores sugerem que aquela reação é uma resposta inflamatória normal a um tecido lesado, envolvida no processo geral de reparação e reconstrução tecidular, rejeitando a hipótese de existir um fenómeno de autoimunidade na regressão espontânea das HDIV73,75.
b. Imunidade Adquirida: perspetiva autoimune
O reconhecimento de antigénios por células apresentadoras de antigénios (APC) representa um passo importante no despoletar de uma resposta imune adaptativa. Após a captura do antigénio, as APC migram para os tecidos linfoides periféricos apresentando o antigénio aos linfócitos T CD4+ naïve19,76. O NP herniado parece representar o local onde os antigénios são expostos ao sistema imune. Deste modo, dado o DIV ser avascular, apenas quando ele é exposto à circulação sanguínea e, por sua vez, ao sistema imune, é que os antígenos derivados do NP podem migrar para os tecidos linfoides e sofrer a tolerância imunológica77. Quando esta tolerância imunológica não ocorre, é iniciada uma resposta imune adaptativa mediada por células. Os macrófagos e as células dendríticas são exemplos de APC19,76.
Geiss et al. observou que nos infiltrados de sequestros de HDIV as células dendríticas plasmocitóides (PDC) e os linfócitos T de memória eram detetados em proporções significativamente superiores em relação aos macrófagos, revelando a predominância das PDC neste tipo de hérnia. Concluindo, estas descobertas são indicativas que as PDC estão envolvidas na iniciação de uma resposta autoimune específica direcionada ao NP herniado, enquanto os macrófagos, além de reforçarem esta resposta pelas suas propriedades enquanto APC, medeiam a reabsorção do DIV através das suas propriedades fagocíticas78.
Em estudos imunohistoquímicos de material herniado proveniente de discectomia, a infiltração de células inflamatórias mostrou ser formada por macrófagos e linfócitos T, sendo estes originários dos vasos sanguíneos presentes na periferia das HDIV9.
O corpo humano encontra-se num balanço imune de linfócitos T efetores, regulando positiva ou negativamente as respostas imunes específicas, respetivamente através de linfócitos T CD4+ e linfócitos T CD8+. O rácio linfócitos T CD4+/CD8+ representa o balanço da imunidade, pelo que qualquer alteração deste corresponde a uma provável desregulação da resposta imune específica19,79,80.
Tian et al. através da realização de citometria de fluxo do sangue periférico colhido em doentes com HDIV, observou que ocorreria um aumento significativo da quantidade de linfócitos T CD4+, assim como o aumento do rácio de linfócitos T CD4+/CD8+, acompanhados de uma diminuição da quantidade de linfócitos T CD8+, em comparação a doentes sem HDIV79,80. No caso dos linfócitos T CD4+ e do rácio linfócitos T CD4+/CD8+ a sua quantidade era significativamente superior nas HDIVE em relação às HDIVC e aos controlos, respetivamente79. Estas evidências indicam que ocorrem alterações no sistema imune após a HDIV, principalmente quando ocorre extrusão do conteúdo herniado através do LLP, apoiando a hipótese relacionada com a resposta autoimune específica79,80.
A HDIV ao sofrer extrusão é invadida por células inflamatórias, que através da libertação de citocinas e quimiocinas, parecem promover uma resposta Th1 com supressão da resposta Th221. O resultado é a violação do estado imunoprivilegiado ocorrendo uma resposta inflamatória por reação imune direta ao DIV herniado21.
Os linfócitos Th1 secretam altos níveis de IL-2, TNFα e IFNγ que ativam macrófagos e promovem respostas celulares imunologicamente mediadas. Os linfócitos Th2 produzem uma variedade de citocinas anti-inflamatórias, incluindo IL-4, IL-5, IL-10 e IL-1369.
A IL-4 é capaz de suprimir citocinas derivadas dos monócitos, incluindo TNFα, IL-1ß, IL-6, CXCL8 e a CCL3. Podemos observar que as protusões do DIV se relacionam com um predomínio da concentração sérica IL-4, enquanto as extrusões do DIV com o predomínio de TNFα81. Localmente, as concentrações de IFNγ e IL-12 foram superiores nas HDIVE, ao contrário da IL-4, em que a sua concentração foi superior nas HDIVC21. A positividade para o IFNγ, a presença de macrófagos e o aumento de celularidade no DIV parece traduzir resposta do tipo Th1 na HDIV82. A análise do plasma sérico demonstrou significativamente um aumento dos níveis de fatores pro inflamatórios (IL-1ß, IL-2, IL-12, IFNγ e TNFα) e uma diminuição dos níveis de IL-4 em indivíduos com evidência de HDIV, em comparação aos indivíduos sem a patologia. Apesar de numerosos fatores afetarem os níveis no sangue periférico, uma total avaliação do sistema imune poderá ser útil na definição do tratamento médico na HDIV69. Uma nota final para o papel da citocina proinflamatória IL-2, que está aumentada em HDIV, possuindo um papel patogénico no prolapso de DIV, nomeadamente através da promoção da apoptose celular e da degradação da matriz extracelular no NP de DIV83.
Num outro estudo, Geiss et al. verificou que o NP autólogo transplantado subcutaneamente atrai uma proporção significativamente elevada de linfócitos T (com maior proporção de linfócitos T CD4+ em comparação aos linfócitos T CD8+) e linfócitos B ativados expressando a imunoglobulina kappa. Observaram também uma grande quantidade de células T CD45RC imunorreativas, indicando a presença de linfócitos T de memória. A grande expressão de linfócitos B expressando cadeias leves kappa de IgG aponta mais para a presença de linfócitos B de memória, do que de plasmócitos produtores de autoanticorpos. Este estudo permitiu confirmar que o NP herniado atrai linfócitos T e B, mimetizando a sua exposição aquando da herniação do DIV, e mostrando a sua potencialidade para gerar uma reposta imune específica devido às suas propriedades autoimunes. Os linfócitos T CD4+ e linfócitos B devem estar envolvidos nesta resposta84, pelo que a sua prevalência é superior nos sequestros comparativamente às extrusões72.
A ativação de linfócitos T representa o marco na geração de uma possível resposta autoimune, levando subsequentemente à diferenciação de linfócitos B84.
A quantidade de linfócitos T e macrófagos, assim como as concentrações de imunoglobulinas IgM e IgG no sangue periférico, foram superiores em pacientes com HDIVE comparativamente àqueles com protusões79.
Satoh et al. verificou através de estudos imunohistoquímicos do conteúdo de tecido herniado a existência de complexos antigénio-anticorpo localizados na cápsula pericelular dos condrócitos da HDIV. Tais complexos não foram observados nas amostras de NP não herniado dos mesmos doentes e no grupo de controlo. Os autores sugerem que a IgG é o anticorpo primário, ligando-se a estruturas específicas no tecido da HDIV. Contudo, afirmam que o antigénio responsável é desconhecido85.
Verificou-se que a IgM e a IgG são depositadas em redor dos vasos recém-formados e do NP das HDIV, concluindo que estas imunoglobulinas possuem um papel importante na resposta imune do HDIV79.
7. Fagocitose
Independentemente do processo imune subjacente, é de algum modo consensual que a reabsorção espontânea ocorre através da fagocitose do tecido herniado, mediada pelos macrófagos61,62,65,86,87. As principais ações mediadas pelos macrófagos parecem decorrer da sua poderosa atividade fagocítica e secretora de enzimas com um papel na digestão e degradação do material herniado62.
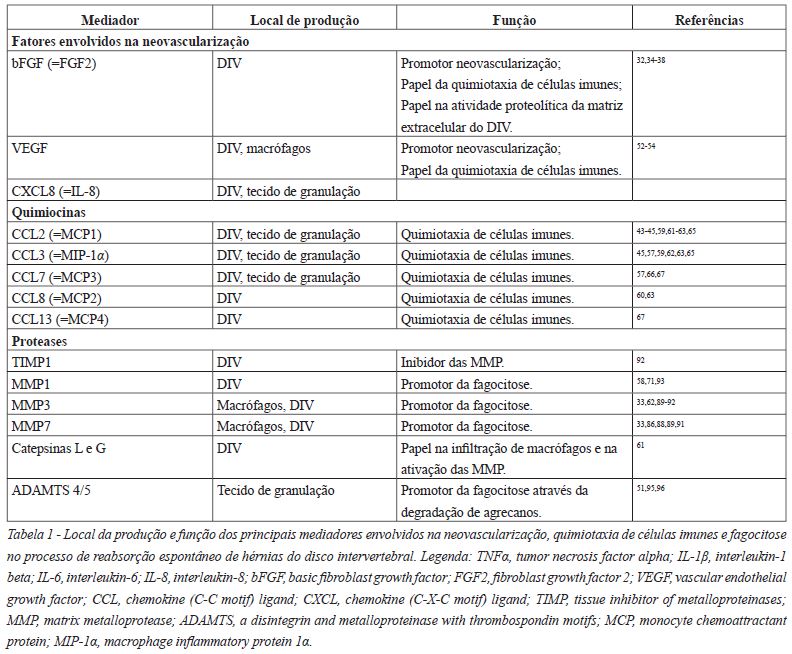
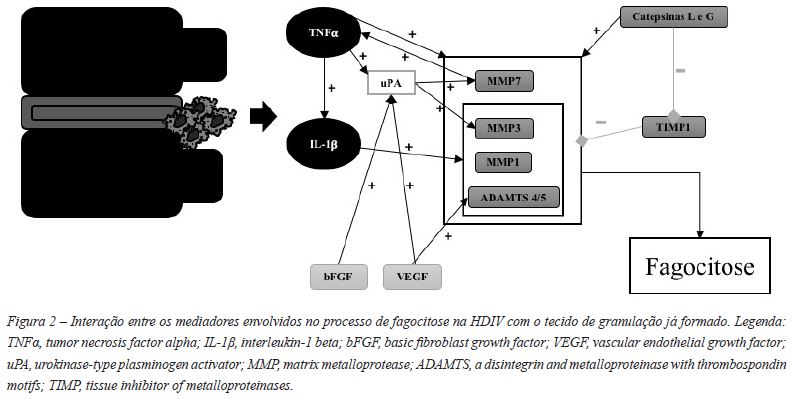
Ikeda et al. verificou a existência de proteínases em redor dos infiltrados macrófagicos54. Estas enzimas, maioritariamente metaloproteínases da matriz (MMP), ADAMTS ou catepsinas, estão envolvidas na degradação da matriz extracelular do NP através do seu papel na decomposição de colagénio, agrecanos e proteoglicanos54,55,61,71.
As quimiocinas além de influenciarem a infiltração de macrófagos, parecem também promover a ativação das MMP33,43,59,61,62,65-67. A produção MMP é regulada por inibidores tecidulares da metaloproteínase 1 (TIMP1), glicoproteínas naturalmente produzidas no DIV que inibem as MMP.
De um modo geral os macrófagos expressam caracteristicamente MMP733,86,88,89 e MMP333,90. Por sua vez, as células do DIV expressam caracteristicamente MMP333,89,91,92, TIMP192, MMP786,91 e MMP193 (Tabela 1).
A elevação dos níveis de MMP, principalmente de MMP3, ocorre paralelamente ao aumento do número de macrófagos33,61,87. As MMP1 e MMP3 estão envolvidas na formação do tecido de granulação, na degradação e na reabsorção dos tecidos da HDIV71 (Figura 2).
No tecido do DIV saudável a expressão de MMP1 é superior à de MMP3, contudo diminui com a evolução da degeneração do DIV, tornando-se os níveis de ambas as MMP similares num estado avançado da degeneração. Quando ocorre herniação do DIV os níveis de MMP3 tornam-se superiores aos de MMP1, indicando que a reabsorção do tecido herniado parece estar mais dependente da MMP393. A produção de MMP158, MMP358,90,92 e MMP786 aumentam significativamente quando o NP herniado é exposto ao espaço epidural, estando aumentadas nas HDIVE em comparação às HDIVC.
Haro et al. através de um modelo de cocultura de macrófagos de ratinhos com tecidos do DIV, relacionou a expressão de MMP7, MMP3 e TNFα pelos macrófagos com a redução do peso da amostra de tecido do DIV e a sua reabsorção. Concluiu que a produção de MMP7 por macrófagos é requerida para a indução de MMP3 no DIV e este efeito é mediado por uma via de sinalização que envolve o TNFα. O efeito da MMP7 não foi direto na síntese de TNFα, mas na sua libertação na superfície celular88. Temporalmente ocorre primeiro a expressão de TNFα, seguindo-se a expressão de uPA, de MMP7 e de MMP3. A uPA ativa as formas latentes de MMP3 e MMP789. O TNFα é assim necessário, mas não suficiente, para a reabsorção do DIV88.
O TNFα induz a produção de MMP3 nas células do DIV de forma significativa58,88,91,94 comparativamente, a IL-1ß apresenta um efeito mais modesto na indução da MMP3, sendo esta mais responsiva ao TNFα88,94.
Quando estimuladas com IL-1ß e TNFα, as HDIVE produzem uma maior quantidade de MMP1 e MMP3 do que as HDIVC58. Os macrófagos produzem TNFα e MMP7, resultando na libertação de mais TNFα88. Ocorre assim a consequente produção de MMP3 pelas células do DIV que contribui para a geração de fatores promotores da migração de macrófagos, através do seu papel na comunicação condrócito/macrófago33,88,91. O papel da expressão de MMP3 na degradação dos diversos componentes da matriz do DIV94 parece estar mais relacionado com a mediação da interação célula-célula do que com a própria degradação da matriz dos tecidos herniados33. A MMP7 apesar de ser necessária para a libertação de TNFα e, indiretamente, para a infiltração de macrófagos no tecido do DIV88, é uma enzima potente envolvida na degradação direta dos tecidos de HDIV. Neste sentido, foi demonstrado que a MMP7 leva uma diminuição significativa, enquanto a MMP3 origina apenas uma diminuição relativa do peso das amostras no modelo estudado70. A indução da MMP3 nas células do DIV foi necessária, mas não suficiente, para ocorrer degradação da matriz no modelo de reabsorção da HDIV33.
Os autores especulam que as várias famílias de proteínases atuem em conjunto ao mediar a destruição da cartilagem no processo de reabsorção do HDIV86,88.
As catepsinas são proteínases de serina produzidas pelas células do DIV independentemente da infiltração de macrófagos, ao contrário das MMP. O pico da quantidade de catepsinas G e L ocorre antes da invasão do DIV por macrófagos, sendo o pico da catepsina L mais tardio do que o da catepsina G. A catepsina G e a CCL2(=MCP1) parecem ativar-se antes do início do processo de reabsorção dependente dos macrófagos. Estas parecem influenciar a própria infiltração de macrófagos e a ativação das MMP produzidas por eles mesmo. Efetivamente, estas catepsinas ativam as MMP latentes e degradam TIMP. A localização da catepsina G e das MMP foi semelhante, o que reforça
esta evidência61.
As ADAMTS4/5 são importantes enzimas envolvida na degradação de agrecanos51,95,96. Os macrófagos dos tecidos de granulação expressam ADAMTS4, sugerindo o seu envolvimento na regressão da HDIV. A contagem de macrófagos e a expressão de ADAMTS4 foi significativamente superior nas HDIVE, comparando com as HDIVC, pelo que estas evidências poderão explicar a razão pela qual as primeiras são mais rapidamente reabsorvidas51. Tanto a ADAMTS4, como a ADAMTS5 são necessárias para a degradação dependente de citocinas nas células do NP humano, pelo que a sua função não parece ser redundante, sugerindo o seu papel em patologias do DIV96.
CONCLUSÃO
Ainda existe controvérsia em relação ao tipo de imunidade implicada na reabsorção das HDIV, ficando os autores com a impressão que quer a imunidade inata, quer a imunidade específica podem estar ambas implicadas. É de algum modo consensual que no processo de reabsorção das HDIV estão envolvidos fenómenos de neovascularização, quimiotaxia de células imunes e formação de um tecido de granulação, culminando na fagocitose onde várias famílias de proteínases atuam em conjunto para destruir o material herniado.
Foram encontrados fatores que se associam a uma maior probabilidade de reabsorção como as HDIVE, o maior volume e intensidade de sinal do material herniado, enquanto que por outro lado um maior conteúdo de plataforma cartilagínea e a associação a alterações Modic influenciam negativamente aquele processo.
REFERÊNCIAS BIBLIOGRÁFICAS
1. Hoy D. A systematic review of the global prevalence of low back pain. Arthritis Rheum. 2012 Jun; 64 (6): 2028-2037
2. Walker BF. The prevalence of low back pain: a systematic review of the literature from 1966 to 1998. J Spinal Disord. 2000 Jun; 13 (3): 205-217
3. Simon J. Discogenic low back pain. Phys Med Rehabil Clin N Am. 2014; 25 (2): 305-317 [ Links ]
4. Bozzao A. Lumbar disk herniation: MR imaging assessment of natural history in patients treated without surgery. Radiology. 1992 Sep; 185 (1): 135-141
5. Komori H. The natural history of herniated nucleus pulposus with radiculopathy. Spine (Phila Pa 1976). 1996 Jan 15; 21 (2): 225-229
6. Macki M. Spontaneous regression of sequestrated lumbar disc herniations: Literature review. Clin Neurol Neurosurg. 2014 May; 120: 136-141
7. Zhong M. Incidence of Spontaneous Resorption of Lumbar Disc Herniation: A Meta-Analysis. Pain Physician. 2017 Jan; 20 (1): 45-52
8. Buckwalter JA. Aging and degeneration of the human intervertebral disc. Spine (Phila Pa 1976). 1995 Jun 1; 20 (11): 1307-1314
9. Arai Y. Immunohistological study of intervertebral disc herniation of lumbar spine. J Orthop Sci. 2000; 5 (3): 229-231 [ Links ]
10. Molinos M. Inflammation in intervertebral disc degeneration and regeneration. J R Soc Interface. 2015 Mar 6; 12 (104): 20141191
11. Slavin KV. Spontaneous regression of a large lumbar disc herniation: report of an illustrative case. Surg Neurol. 2001 Nov; 56 (5): 333-336
12. Henmi T. Natural history of extruded lumbar intervertebral disc herniation. J Med Invest. 2002 Fev; 49 (1-2): 40-43
13. Citisli V, Ibrahimoglu M. Spontaneous remission of a big subligamentous extruded disc herniation: case report and review of the literature. Korean J Spine. 2015 Mar; 12 (1): 19-21
14. Splendiani A. Spontaneous resolution of lumbar disk herniation: predictive signs for prognostic evaluation. Neuroradiology. 2004 Nov; 46 (11): 916-922
15. Sari H. Computed tomographic evaluation of lumbar spinal structures during traction. Physiother Theory Pract. 2005 Jan; 21 (1): 3-11
16. Ito T. Histologic evidence of absorption of sequestration-type herniated disc. Spine (Phila Pa 1976). 1996 Jan 15; 21 (2): 230-234
17. Liu ZH. FasL expression on human nucleus pulposus cells contributes to the immune privilege of intervertebral disc by interacting with immunocytes. Int J Med Sci. 2013 Jun 21; 10 (8): 1053-1060
18. Sun Z. FasL on human nucleus pulposus cells prevents angiogenesis in the disc by inducing Fasmediated apoptosis of vascular endothelial cells. Int J Clin Exp Pathol. 2013 Oct 15; 6 (11): 2376-2385
19. Abbas A, Lichtman A, Pillai S. Cellular and Molecular Immunology. 9th. Philadelphia: Elsevier; 2018. [ Links ]
20. Park JB, Chang H, Kim KW. Expression of Fas ligand and apoptosis of disc cells in herniated lumbar disc tissue. Expression of Fas ligand and apoptosis of disc cells in herniated lumbar disc tissue. 2001 Mar 15; 26 (6): 618-621
21. Park JB, Chang H, Kim YS. The pattern of interleukin-12 and T-helper types 1 and 2 cytokine expression in herniated lumbar disc tissue. Spine (Phila Pa 1976). 2002 Oct 1; 27 (19): 2125-2128
22. Nerlich AG. Immunolocalization of phagocytic cells in normal and degenerated intervertebral discs. Spine (Phila Pa 1976). 2002 Nov 15; 27 (22): 2484-2490
23. Chen YF. Insights into the hallmarks of human nucleus pulposus cells with particular reference to cell viability, phagocytic potential and long process formation. Int J Med Sci. 2013 Oct 31; 10 (13): 1805-1816
24. Rade M. Vertebral Endplate Defect as Initiating Factor in Intervertebral Disc Degeneration; Strong Association between Endplate Defect and Disc Degeneration in the General Population. Spine (Phila Pa 1976). 2018 Mar 15; 43 (6): 412-419
25. Rajasekaran S. ISSLS Prize winner: The anatomy of failure in lumbar disc herniation: an in vivo, multimodal, prospective study of 181 subjects. Spine (Phila Pa 1976). 2013 Aug 1; 38 (17): 1491-1500
26. Sahoo MM. Significance of Vertebral Endplate Failure in Symptomatic Lumbar Disc Herniation. Global Spine J. 2017 May; 7 (3): 230-238
27. Ahn SH, Ahn MW, Byun WM. Effect of the transligamentous extension of lumbar disc herniations on their regression and the clinical outcome of sciatica. Spine (Phila Pa 1976). 2000 Fev 15; 25 (4): 475-480
28. Seo JY. Three-dimensional analysis of volumetric changes in herniated discs of the lumbar spine: does spontaneous resorption of herniated discs always occur. Spine J. 2016 May; 25 (5): 1393-1402
29. Autio RA. Determinants of spontaneous resorption of intervertebral disc herniations. Spine (Phila Pa 1976). 2006 May 15; 31 (11): 1247-1252
30. Ohba T. TNF-alpha-induced NF-kappaB signaling reverses age-related declines in VEGF induction and angiogenic activity in intervertebral disc tissues. J Orthop Res. 2009 Fev; 27 (2): 229-235
31. Binch AL. Expression and regulation of neurotrophic and angiogenic factors during human intervertebral disc degeneration. Arthritis Res Ther. 2014; 16 (5): 416 [ Links ]
32. Doita M. Immunohistologic study of the ruptured intervertebral disc of the lumbar spine. Spine (Phila Pa 1976). 1996 Jan 15; 21 (2): 235-241
33. Haro H. Matrix metalloproteinase-3-dependent generation of a macrophage chemoattractant in a model of herniated disc resorption. J Clin Invest. 2000 Jan; 105 (2): 133-141
34. Li X. Action of fibroblast growth factor-2 on the intervertebral disc. Arthritis Res Ther. 2008; 10 (2): 48 [ Links ]
35. Tolonen J. Basic fibroblast growth factor immunoreactivity in blood vessels and cells of disc herniations. Spine (Phila Pa 1976). 1995 Fev 1; 20 (3): 271-276
36. Melrose J. Increased nerve and blood vessel ingrowth associated with proteoglycan depletion in an ovine anular lesion model of experimental disc degeneration. Spine (Phila Pa 1976). 2002 Jun 15; 27 (12): 1278-1285
37. Minamide A. Effects of basic fibroblast growth factor on spontaneous resorption of herniated intervertebral discs. An experimental study in the rabbit. Spine (Phila Pa 1976). 1999 May 15; 24 (10): 940-945
38. Montesano R. Basic fibroblast growth factor induces angiogenesis in vitro. Proc Natl Acad Sci U S A. 1986 Oct; 83 (19): 7297-7301
39. Tsarouhas A. Molecular profile of major growth factors in lumbar intervertebral disc herniation:Correlation with patient clinical and epidemiological characteristics. Mol Med Rep. 2017 Apr; 15 (4): 2195-2203
40. Haro H. Vascular endothelial growth factor (VEGF)-induced angiogenesis in herniated disc resorption. J Orthop Res. 2002 May; 20 (3): 409-415
41. Koike Y. Angiogenesis and inflammatory cell infiltration in lumbar disc herniation. Spine (Phila Pa 1976). 2003 Sep 1; 28 (17): 1928-1933
42. Deng X. Elevated interleukin-6 expression levels are associated with intervertebral disc degeneration. Exp Ther Med. 2016 Apr; 11 (4): 1425-1432
43. Walter BA. Inflammatory Kinetics and Efficacy of Anti-inflammatory Treatments on Human Nucleus Pulposus Cells. Spine (Phila Pa 1976). 2015 Jul 1; 40 (13): 955-963
44. Takada T. Intervertebral disc and macrophage interaction induces mechanical hyperalgesia and cytokine production in a herniated disc model in rats. Arthritis Rheum. 2012 Aug; 64 (8): 2601-2610
45. Burke JG. Spontaneous production of monocyte chemoattractant protein-1 and interleukin-8 by the human lumbar intervertebral disc. Spine (Phila Pa 1976). 2002 Jul 1; 27 (13): 1402-1407
46. Hirsch C, Schajowicz F. Studies on structural changes in the lumbar annulus fibrosus. Acta Orthop Scand. 1952; 22 (1-4): 184-231 [ Links ]
47. Roberts S. Histology and pathology of the human intervertebral disc. J Bone Joint Surg Am. 2006 Apr; 88 (2): 10-14
48. Shan Z. Spontaneous resorption of lumbar disc herniation is less likely when modic changes are present. Spine (Phila Pa 1976). 2014 Apr 20; 39 (9): 736-744
49. Carreon LY. Neovascularization induced by anulus and its inhibition by cartilage endplate. Its role in disc absorption. Spine (Phila Pa 1976). 1997 Jul 1; 22 (13): 1429-1434
50. Modic MT. Degenerative disk disease:assessment of changes in vertebral body marrow with MR imaging. Radiology. 1988 Jan; 166 (1 Pt 1): 193-199
51. Hatano E. Expression of ADAMTS-4 (aggrecanase-1) and possible involvement in regression of lumbar disc herniation. Spine (Phila Pa 1976). 2006 Jun 1; 31 (13): 1426-1432
52. Olmarker K. Neovascularization and neoinnervation of subcutaneously placed nucleus pulposus and the inhibitory effects of certain drugs. Spine (Phila Pa 1976). 2005 Jul 1; 30 (13): 1501-1504
53. Ratsep T, Minajeva A, Asser T. Relationship between neovascularization and degenerative changes in herniated lumbar intervertebral discs. Eur Spine J. 2013 Nov; 22 (11): 2474-2480
54. Ikeda T. Pathomechanism of spontaneous regression of the herniated lumbar disc: histologic and immunohistochemical study. J Spinal Disord. 1996; 9 (2): 136-140 [ Links ]
55. Kobayashi S. Ultrastructural analysis on lumbar disc herniation using surgical specimens: role of neovascularization and macrophages in hernias. Spine (Phila Pa 1976). 2009 Apr 1; 34 (7): 655-662
56. Ozaki S. Neovascularization of the outermost area of herniated lumbar intervertebral discs. J Orthop Sci. 1999; 4 (4): 286-292 [ Links ]
57. Haro H. Upregulated expression of chemokines in herniated nucleus pulposus resorption. Spine (Phila Pa 1976). 1996 Jul 15; 21 (14): 1647-1652
58. Doita M. Influence of macrophage infiltration of herniated disc tissue on the production of matrix metalloproteinases leading to disc resorption. Spine (Phila Pa 1976). 2001 Jul 15; 26 (14): 1522-1527
59. Kikuchi T. Monocyte chemoattractant protein-1 in the intervertebral disc. A histologic experimental model. Spine (Phila Pa 1976). 1998 May 15; 23 (10): 1091-1099
60. Kawaguchi S. Chemokine profile of herniated intervertebral discs infiltrated with monocytes and macrophages. Spine (Phila Pa 1976). 2002 Jul 15; 27 (14): 1511-1516
61. Meng W. Localization of cathepsins G and L in spontaneous resorption of intervertebral discs in a rat experimental model. J Musculoskelet Neuronal Interact. 2001 Dec; 2 (2): 171-176
62. Yoshida M. Intervertebral disc cells produce tumor necrosis factor alpha, interleukin-1beta, and monocyte chemoattractant protein-1 immediately after herniation: an experimental study using a new hernia model. Spine (Phila Pa 1976). 2005 Jan 1; 30 (1): 55-61
63. Phillips KL. The cytokine and chemokine expression profile of nucleus pulposus cells: implications for degeneration and regeneration of the intervertebral disc. Arthritis Res Ther. 2013; 15 (6): 213 [ Links ]
64. Huang KY. IL-20 may contribute to the pathogenesis of human intervertebral disc herniation. Spine (Phila Pa 1976). 2008 Aug 1; 33 (19): 2034-2040
65. Haro H. Sequential dynamics of monocyte chemotactic protein-1 expression in herniated nucleus pulposus resorption. J Orthop Res. 1997 Aug; 15 (5): 734-741
66. Wang SL. Effects of TGF-beta1 and IL-1beta on expression of ADAMTS enzymes and TIMP-3 in human intervertebral disc degeneration. Exp Ther Med. 2013 Dec; 6 (6): 1522-1526
67. Moen A. Inflammatory Serum Protein Profiling of Patients with Lumbar Radicular Pain One Year after Disc Herniation. Int J Inflam. 2016; (2016): 3874964 [ Links ]
68. Bobechko WP, Hirsch C. Auto-Immune Response to Nucleus Pulposus in the Rabbit. J Bone Joint Surg Br. 1965 Aug; 47: 574-580
69. Akyol S, Hanci M. TH1 and TH2 Cytokines Production and NK Cell Level Assessment in Peripheral Blood of Patients with DDH. Indian J Surg. 2013 Aug; 75 (4): 294-297
70. Haro H. Experimental studies on the effects of recombinant human matrix metalloproteinases on herniated disc tissues--how to facilitate the natural resorption process of herniated discs. J Orthop Res. 2005 Mar; 23 (2): 412-419
71. Matsui Y. The involvement of matrix metalloproteinases and inflammation in lumbar disc herniation. Spine (Phila Pa 1976). 1998 Apr 15; 23 (8): 863-868
72. Habtemariam A. Inflammatory cells in fullthickness anulus injury in pigs. An experimental disc herniation animal model. Spine (Phila Pa 1976). 1998; 23 (5): 524-529 [ Links ]
73. Kawaguchi S. Immunophenotypic analysis of the inflammatory infiltrates in herniated intervertebral discs. Spine (Phila Pa 1976). 2001 Jun 1; 26 (11): 1209-1214
74. Murai K. Primary immune system responders to nucleus pulposus cells: evidence for immune response in disc herniation. Eur Cell Mater. 2010 Jan 14; 19: 13-21
75. Zhou G. Effects of human midkine on spontaneous resorption of herniated intervertebral discs. Int Orthop. 2010 Jan; 34 (1): 103-108
76. Villadangos JA, Young L. Antigen-presentation properties of plasmacytoid dendritic cells. Immunity. 2008 Sep 19; 29 (3): 352-361
77. Geiss A. Autologous nucleus pulposus primes T cells to develop into interleukin-4-producing effector cells: an experimental study on the autoimmune properties of nucleus pulposus. J Orthop Res. 2009 Jan; 27 (1): 97-103
78. Geiss A. Plasmacytoid dendritic cells and memory T cells infiltrate true sequestrations stronger than subligamentous sequestrations: evidence from flow cytometric analysis of disc infiltrates. Eur Spine J. 2016 May; 25 (5): 1417-1427
79. Ma XL. A study of the relationship between type of lumbar disc herniation, straight leg raising test and peripheral T lymphocytes. Orthop Surg. 2010 Fev; 2 (1): 52-57
80. Tian P. Correlation between radiculalgia and counts of T lymphocyte subsets in the peripheral blood of patients with lumbar disc herniation. Orthop Surg. 2009 Nov; 1 (4): 317-321
81. Zu B. Serum Levels of the Inflammatory Cytokines in Patients with Lumbar Radicular Pain Due to Disc Herniation. Asian Spine J. 2016 Oct; 10 (15): 843-849
82. Shamji MF. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthritis Rheum. 2010 Jul; 62 (7): 1974-1982
83. Wang Z. Interleukin-2 is upregulated in patients with a prolapsed lumbar intervertebral disc and modulates cell proliferation, apoptosis and extracellular matrix metabolism of human nucleus pulposus cells. Exp Ther Med. 2015 Dec; 10 (6): 2437-2443
84. Geiss A. Autoimmune properties of nucleus pulposus: an experimental study in pigs. Spine (Phila Pa 1976). 2007 Jan 15; 32 (2): 168-173
85. Satoh K. Presence and distribution of antigenantibody complexes in the herniated nucleus pulposus. Spine (Phila Pa 1976). 1999 Oct 1; 24 (19): 1980-1984
86. Haro H. Up-regulated expression of matrilysin and neutrophil collagenase in human herniated discs. J Spinal Disord. 1999 Jun; 12 (3): 245-249
87. Weiler C. 2002 SSE Award Competition in Basic Science: expression of major matrix metalloproteinases is associated with intervertebral disc degradation and resorption. Eur Spine J. 2002 Aug; 11 (4): 308-320
88. Haro H. Matrix metalloproteinase-7-dependent release of tumor necrosis factor-alpha in a model of herniated disc resorption. J Clin Invest. 2001 Jan; 105 (2): 143-150
89. Kato T. Sequential dynamics of inflammatory cytokine, angiogenesis inducing factor and matrix degrading enzymes during spontaneous resorption of the herniated disc. J Orthop Res. 2004; 22 (4): 895-900 [ Links ]
90. Haro H. Chemonucleolysis with human stromelysin-1. Spine (Phila Pa 1976). 1997 May 15; 22 (10): 1098-1104
91. Bachmeier BE. Matrix metalloproteinase expression levels suggest distinct enzyme roles during lumbar disc herniation and degeneration. Eur Spine J. 2009 Nov; 18 (11): 1573-1586
92. Tsuru M. Electron microscopic observation of established chondrocytes derived from human intervertebral disc hernia (KTN-1) and role of macrophages in spontaneous regression of degenerated tissues. Spine J. 2001 Dec; 1 (6): 422-431
93. Genevay S. Influence of cytokine inhibitors on concentration and activity of MMP-1 and MMP-3 in disc herniation. Arthritis Res Ther. 2009; 11 (6): 169 [ Links ]
94. Wang X. Tumor necrosis factoralpha-and interleukin-1beta-dependent matrix metalloproteinase-3 expression in nucleus pulposus cells requires cooperative signaling via syndecan 4 and mitogen-activated protein kinase-NF-kappaB axis: implications in inflammatory disc disease. Am J Pathol. 2014 Sep; 184 (9): 2560-2572
95. Sun Z. IL-1beta promotes ADAMTS enzymemediated aggrecan degradation through NF-kappaB in human intervertebral disc. J Orthop Surg Res. 2015 Oct 6; 10: 159
96. Tian Y. Inflammatory cytokines associated with degenerative disc disease control aggrecanase-1 (ADAMTS-4) expression in nucleus pulposus cells through MAPK and NF-kappaB. Am J Pathol. 2013 Jun; 182 (6): 2310-2321
Conflito de interesse:
Nada a declarar
Belmiro Alves
Rua Adelaide Estrada, 129 - 1º B
4250-016 Ramalde, Porto
Telefone: 91 302 29 55
belmiroliveiralves@gmail.com
Data de Submissão: 2018-06-15
Data de Revisão: 2018-07-01
Data de Aceitação: 2018-07-01


{kind=link}
{kind=link}
{kind=link}