










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkArquivos de Medicina
versão On-line ISSN 2183-2447
Arq Med vol.28 no.6 Porto dez. 2014
ARTIGO DE REVISÃO
Dominância e bonança: mapeando e integrando a psicobiologia e a ecologia dos determinantes da razão sexual primária humana
Dominance and lull: mapping and integrating psychobiological and the ecological determinants of primary sexual ration in humans
Álvaro Machado Dias1,2
1Laboratório de Neurociências Clínicas da Universidade Federal de São Paulo, Brasil
2 Laboratório de Neuroimagem, Instituto de Psiquiatria da Universidade de São Paulo, Brasil
RESUMO
Contexto: Até pouco tempo atrás, acreditava-se que a razão sexual primária (quantidade de nascidos do sexo masc./fem.) fosse um fenômeno estocástico, ainda que achados anedóticos, como alterações no início de guerras e ao longo de períodos caracterizados por catástrofes naturais, sugerissem o contrário. Recentemente, estes achados foram sendo generalizados e hipóteses diversas foram sendo levantadas.
Objetivo: Este estudo tem por objetivos 1. Revisar e categorizar os diferentes planos de desenvolvimento das causas proximais do fenômeno, delimitando os principais fatores não estocásticos em um algoritmo; 2. Apresentar a cascata bioquímica mais pertinentemente implicada; 3. Minerar a literatura em razão sexual envolvendo o principal hormônio implicado no fenômeno das alterações não estocásticas.
Métodos: Todos os estudos de interesse no tema foram compilados e cuidadosamente discutidos nas seções de revisão do presente artigo, do que emergiu o algoritmo inicial. A cascata bioquímica pautou-se por sugestões da literatura e utilizou o KEGG (http://www.genome.jp/kegg/tool/search_pathway.html). A mineração de dados utilizou o Omniviz e dados compilados do PubMed.
Resultados: A razão sexual é profundamente afetada por fatores psicológicos e parece envolver primariamente a quantidade de testosterona materna. A literatura no assunto é escassa e aponta primariamente para estudos em animais.
Palavras-chave: razão sexual, psico-endocrinologia, bioinformática, data mining, evolução
ABSTRACT
Context: until recently, it was believed that the primary sexual reason (masc./fem. offspring) was a stochastic phenomenon, despite anecdotic findings, like alterations in the beginning of wars and in periods of natural catastrophe, pointed in the opposite direction. Recently, these findings were generalized and several hypotheses were raised.
Objective: This study aims to 1. Review and categorize the different proximal level causal mechanisms involved in the phenomenon and incorporate the main non-stochastic factors in an algorithm; 2. Present the most prominent biochemical cascade involved; Mine the literature on sexual reason and the most important hormone involved in non-stochastic effects.
Methods: All studies of interest on this matter were compiled and carefully discussed in the reviewing sections of the paper, leading to the emergence of the algorithm and the epidemiological equation. The biochemical cascade was based on suggestions of the literature and implemented using KEGG (http://www.genome.jp/kegg/tool/search_pathway.html). Data mining was based on Omniviz and PubMed records.
Results: sexual reason is profoundly affected by psychological and seems to primarily involve the amount of maternal testosterone. The literature on the matter is scarce and mainly reduced to studies on animal.
Key-words: sexual reason, psycho-endocrinology, bioinformatics, data mining, evolution
Introdução
Define-se como razão sexual humana à razão entre indivíduos do sexo masculino e indivíduos do sexo feminino. Quanto mais alta a proporção de machos, maior a razão sexual e vice e versa. Está bastante consolidado que ela varia ao longo das diferentes faixas etárias, apresentando uma curva de desenvolvimento decrescente que acompanha o envelhecimento (1.1=x=0.9), refletindo o aumento proporcional do número de mulheres, o que pode ser associado à maior suscetibilidade masculina a vários insultos ambientais e maus funcionamentos orgânicos (Kraemer, 2000).
A razão sexual pode ser dividida por faixas etárias, sendo assim considerada primária (concepção), secundária (nascimento) e terciária (ao longo da vida, após o nascimento); ela também se diferencia em física (total) e operacional (relativa aos indivíduos em idade reprodutiva).
Diversas variáveis endógenas e ecológicas estão relacionadas a alterações pontuais deste índice, as quais podem ter papel fundamental no planejamento estratégico de caráter demográfico. Dito isto, é importante ter em vista que desequilíbrios proporcionais entre os sexos não tendem a se manter indefinidamente, posto que a maior disponibilidade de um sexo leva ao aumento da disputa pelo outro, como argumentado por Wilson (1978).
Alterações da razão sexual terciária naturalmente incorporam os efeitos das secundárias e primárias e assim respectivamente. Neste sentido, iniciamos esta empreitada pela sugestão de utilizar a nomenclatura alterações primárias, secundárias e terciárias para nos referirmos ao momento de ocorrência dos eventos críticos, ao invés de se olhar para o momento do recorte.
Demograficamente, as variações terciárias são as mais conhecidas. Por exemplo, sendo a maioria dos combatentes do sexo masculino, decorre o fato de que guerras prolongadas costumam afetar a razão sexual da população, o mesmo se aplicando a al-guns movimentos migratórios.
Alterações secundárias são em grande medida relacionadas a avanços no índice de desenvolvimento humano (IDH), [(l + E + R)/3] (ONU, 2009), sob o mote de que os fetos do sexo masculino são mais suscetíveis a instabilidades do desenvolvimento (Ulizzi, 1993), beneficiando-se proporcionalmente mais de melhorias nutricionais, saneamento básico e acompanhamento obstetrício (note: l = longevidade, E = educação, R = renda).
A este fato se associa à perspectiva de que gestações de fetos masculinos dão origem a maior número de partos prematuros, cesarianas, além de implicar em maior risco de insucesso no trabalho de parto e outras condições clínicas indiretas, como a diabetes mellitus gestacional (para um estudo clínico ver: Joseph, et al., 2005; para uma revisão cobrindo um período de 21 anos: di Renzo, et al., 2007); com base nisto, pode-se dizer que o avanço e popularização de tecnologias médicas não apenas cobre uma demanda alinhada ao estabelecimento da homeostase ecológica, mas efetivamente institui alterações na razão sexual típica.
Secundariamente, despontam as demandas regionais por descendentes de um dos sexos. Sendo a prática de interrupção deliberada de gestações com vias à determinação sexual, consideravelmente frequente em alguns países (Gupta & bhat, 1997; Jha, et al., 2006; Johansson & nygren, 1991; Miller, 1997; Park & cho, 1995; Tuljapurkar, li, & feldman, 1995). como é de se notar, uma grande divisão categórica se interpõe a ambas as variáveis, à luz do conceito de intencionalidade no processo de determinação da razão sexual.
Menos conhecidas – e tanto mais complexas – são as alterações primárias, que por operarem em um contexto de difícil mapeamento, foram menos submetidas ao escrutínio científico. Estas são em grande medida determinadas pelo perfil hormonal parental, através de interações moleculares, cuja elucidação plena ainda vem sendo aguardada.
Por fim, alterações primárias e secundárias frequentemente se sobrepõem sobre as mesmas vias moleculares, exigindo uma abordagem conjunta desde a distinção vias finais e vias intermediárias, isto é, entre as cascatas moleculares que efetiva e diretamente afetam a probabilidade de que a gravidez seja bem sucedida à luz do sexo do feto e aquelas capazes de gerar efeitos indiretos.
sob este mote, apresentamos a seguir o conjunto das variáveis que afetam a razão sexual entre nascidos humanos e um novo modelo de associação aos fatores ecológicos mais fortemente relacionados às mesmas. subsequentemente apresentaremos um novo algoritmo para a caracterização das cascatas moleculares que mais fortemente influenciam a razão sexual primária.
1. Variáveis ecológicas mais fortemente relacionadas às alterações Da razão sexual humana
Quando se fala em alterações primárias e secundárias da razão sexual humana, fala-se naturalmente em alterações não estocásticas. Considerando que o sexo de um feto é definido pela probabilidade de que um óvulo seja fecundado por um espermatozoide portador de um cromossomo Y (androsperma), no contexto da potencial exposição a milhares de espermatozoides e que um sem fim de fatores podem influenciar a probabilidade de aborto natural, emerge o fato de que a circunscrição dos fatores críticos para a razão sexual humana ao nascimento é intrinsecamente depende de comparações entre grupos desde a circunscrição de um número reduzido de variáveis dependentes.
Estas variáveis podem ser representadas em dois níveis: molecular e ecológico, os quais, no âmbito de possíveis representações estruturais do fenômeno, não devem ser vistos como derradeiramente correlatos, mas antes como parcialmente correlatos e fundamentalmente decorrentes, desde a via ecológico - molecular. Por exemplo, considerando que a erupção de vulcões afete a razão sexual humana ao nascimento, é de se ter em vista que isto apenas ocorra conquanto tal altere o comportamento das variáveis envolvidas na probabilidade deque óvulos sejam fertilizados por androspermas e/ou a probabilidade de abortos desproporcionais entre fetos de um dos sexos, do que se pode concluir pelo estabelecimento de uma peculiar equivalência entre os fenômenos, molecular e ecológico, em representações estruturais; paralelamente, as variáveis moleculares envolvidas não surgem desde a erupção de vulcões, mas antes são alteradas por tal, o que por sua vez ocorre no contexto da existência de outros potenciais agentes probabilísticos (tudo o que afete esta via final) seguindo-se, pois, a pertinência da representação do fenômeno ecológico em cascatas de eventos.
Por fim, faz-se importante considerar que, não apenas por uma questão normativa, mas por determinantes ontológicos, desponte a prerrogativa de se elucidar os elos intermediários entre erupções de vulcões e os outros eventos passíveis de afetar o comportamento da via final das razões sexuais primária e secundária, as quais podemos alinhavar sob o conceito de razão sexual humana ao nascimento.
À luz destes princípios, apresentamos a seguir um algoritmo caracterizando o conjunto de variáveis passíveis de afetar a razão sexual humana ao nascimento:
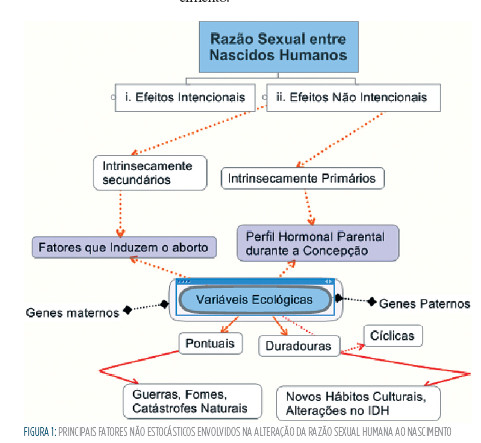
No nosso algoritmo, cores mais vivas representam conceitos e elos mais significativos à compreensão da determinação não estocástica da razão sexual. Conceitos flutuantes (sem caixas) representam variáveis que ao nosso entender são sub-representadas na literatura. Neste sentido (e de maneira inédita) encerraremos esta seção com a apresentação de evidências da hipótese de determinantes genéticos da razão sexual, a qual será apresentada na seção seguinte.
Em termos ecológicos, os efeitos intencionais apresentam-se na forma de abortos induzidos, enquanto os efeitos não intencionais primários e secundários refletem a convergência de variáveis ecológicas comuns.
Frente à necessidade de discriminação destas últimas, introduzimos a seguinte equação: 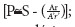 ; onde P =fatores primários, s: fatores secundários, A: número de abortos na amostra, r: razão sexual entre estes, A: número de abortos na média da população, r: razão sexual no âmbito destes. A esta representação acrescentamos ainda o fato de que nem todas as variáveis ecológicas alinhavadas possuem efeitos potencialmente primários e secundários. Quem quiser entender melhor a lógica por trás da equação, pode começar por: http://www.uvm.edu/~aneal/sexratio.html.
; onde P =fatores primários, s: fatores secundários, A: número de abortos na amostra, r: razão sexual entre estes, A: número de abortos na média da população, r: razão sexual no âmbito destes. A esta representação acrescentamos ainda o fato de que nem todas as variáveis ecológicas alinhavadas possuem efeitos potencialmente primários e secundários. Quem quiser entender melhor a lógica por trás da equação, pode começar por: http://www.uvm.edu/~aneal/sexratio.html.
Sob a premissa de que nem sempre se faz possível uma distinção entre primárias e secundárias, apresentamos a seguir as principais variáveis ecológicas e, por fim, genéticas:
a. Variáveis de caráter duradouro, tipo IDH
A razão sexual é sensível ao IDH, apresentando diferenças interpaíses e alterações históricas em diferentes países, correlacionadas ao mesmo.
Um estudo canadense que analisou o censo de nascimentos de 1930-1990 concluiu pela presença de uma redução estatisticamente significativa da razão sexual na porção atlântica do país (p < 0.001), entre 1970-1990 (Allan, brant, seidel, & Jarrell, 1997); um estudo italiano semelhante concluiu pela redução da razão sexual na média dos centros urbanos do país entre 1970-1995 (Astolfi & zonta, 1999), e um estudo dinamarquês encontrou redução da razão sexual no país inteiro, entre as décadas de 1960-1990 (p=0.012) (Møller, 1996).
i. Uma pesquisa escorada em dados compilados pela organização mundial de saúde (WhO), cobrindo o período de 1950-1994 em 29 países, revelou que a razão sexual tende a ser maior nos países europeus centrais e menor nos países asiáticos e na América latina (Parazzini, la vecchia, levi, & franceschi, 1998). Tanto esta quanto a de cima são variáveis fundamentalmente secundárias.
ii. variações individuais de caráter geral: um estudo sugere que condições não ideais para a concepção e gestação diminuam a razão sexual, com destaque para o baixo peso antes da gestação (cagnacci, et. al, 2004). impossível se definir o caráter primário/secundário deste achado.
b. Variáveis de caráter duradouro, tipo mudanças de hábitos culturais
Estas variáveis são significativas, mas não necessariamente capazes de afetar a razão sexual em escala global. É de se ter em vista que também são funções indiretas de alterações no IDH, ainda que não possam ser alinhavadas sob tal eixo.
iii. Efeito de múltiplas gestações: demonstrou-se que uma diminuição da razão sexual secundária em crianças dinamarquesas oriundas de gestações múltiplas (Jacobsen, Moller, & Mouritsen, 1999). O estudo analisou dados de uma população de cerca de 851000 nascidos. Estes resultados podem ser associados ao aumento da prática de reprodução induzida. Trata-se de variável eminentemente primária.
iv. Efeitos da idade parental: alguns estudos sugerem que a razão sexual diminua com a idade da mãe e do pai (respectivamente: Moller, 1996; van der Pal-debruin, Verloove-Vanhorick, &Roeleveld, 1997); segundo uma pesquisa que realizou uma regressão logística para a comparação da importância da idade materna/paterna, é sobretudo a idade da mãe que se faz determinante (Juntunen, Kvist, & Kauppila, 1997). Este fator pode ser relevante para a formação da razão sexual atual, dado o aumento na idade média dos pais que acompanha o aumento da expectativa de vida. Não obstante, este assunto permanece inconcluso, ao passo que alguns auto-res sugerem que tal variável seja estatisticamente desprezível (James, 1987, 2001b). Esta variável pode ser simultaneamente primária (como veremos a seguir) e secundária, dada a chance aumentada de abortos entre mães mais velhas.
c. Variáveis de caráter duradouro, tipo cíclico
As variáveis cíclicas não devem ser levadas em conta para a prospecção de alterações globais da razão sexual humana ao nascimento ao longo da história; por outro lado, são bastante úteis para a investigação das vias finais de tal fenômeno, possuindo caráter simultaneamente primário (como veremos a seguir) e secundário.
v. Efeitos da sazonalidade: um estudo americano do começo da década de 1980 revelou que a razão sexual pode variar em função das estações do ano, simultaneamente em brancos e negros (note que os cruzamentos inter-raciais nesta época eram minoritários), com pico inferior no outono (W. h. James, 1984); este estudo pode ser complementado por outro, que revelou que a razão sexual se acentua na primavera japonesa (Miura, Nonaka, Shimura, & Nakamura, 1983) (março-maio).
Em relação à sibéria (1953-2001), onde as variações de temperatura são extremas, propiciando uma oportunidade significativa para o estudo do fenômeno, sugere-se que o número de nascidos do sexo masculino decresça no último trimestre do ano, eventualmente por alguma influência das temperaturas baixas do primeiro trimestre do ano (Melnikov & Grech, 2003); paralelamente, um estudo cobrindo dados demográficos de 202 países concluiu que a razão sexual é menor nos trópicos do que no ártico (Navara,2009) (dado o qual parece contrastar com o achado siberiano acima), ao passo que uma revisão de literatura argumenta que, de maneira geral, o efeito da sazonalidade não é muito significativo (James, 1996).
d. Variáveis de caráter pontual
Os efeitos destas variáveis são potencialmente primários e secundários, mas há razões para se considerar que os primeiros sejam proeminentes (como veremos a seguir).
vi. Efeitos de guerras: em vários países, verificouse um aumento no número de nascidos do sexo masculino após a segunda guerra mundial (Graffelman & Hoekstra, 2000; Macmahon & Pugh, 1954).
Em relação à guerra Iran-Irque (1980-1988), verificou-se um declínio da razão sexual durante a guerra, mas um aumento subsequente ao seu fim (Ansari-Lari & Saadat, 2002), ao passo que um estudo sobre a guerra da croácia (1991-1995) não encontrou evidências relativas à alteração da razão sexual (Polasek, Kolcic, Kolaric, & Rudan, 2005), o mesmo tenso sido verificado em relação à guerra do Líbano de 2006 (Abu-Musa, usta, Yunis, & Nassar, 2009), cuja duração de apenas 33 dias não pode ser desconsiderada.
vii. Efeitos da exposição a agentes tóxicos: ainda que não haja nenhuma meta-análise no tema, uma visão geral da literatura sugere que a exposição parental a agentes tóxicos a maior responsável por alterações (diminuições) não estocásticas da razão sexual por envenenamento (para estudos experimentais que confirmam isto, ver: Mocarelli, et al., 2000; Ryan, Amirova, & carrier, 2002). Não obstante, isto não é consensual: um estudo sobre exposição à dioxina (2,3,7,8-Tetrachlorodibenzo-pdioxina),emcoletoresdelixojaponeses,revelouleve aumento da razão sexual na progênie destes (Mori, Ogawa, Koda, Kumagai, & ueno, 2004), achado o qual foi contradito por outros estudos (Mocarelli, brambilla, Gerthoux, Jr, & Needham, 1996; Paolo Mocarelli, et al., 2000). Estudo pioneiro, realizado na cidade de São Paulo, com camundongos, revelou uma correlação estatisticamente significativa entre a maior presença de poluentes atmosféricos e me-nor razão sexual, nas diferentes partes da cidade, a qual foi relacionada a alterações morfológicas dos espermatozoides, entre outras (Lichtenfels, 2006).
viii. Efeitos de catástrofes naturais: o terremoto de Kobe (Japão, 1995) foi relacionado a uma diminuição da razão sexual local (Fukuda, Fukuda, Shimizu,&Moller,1998), achado o qual igualmente se verificou no terremoto de Kerman (Iran, 2003) (Saadat, 2008).
e. Efeitos de genes
ix. Não se encontra na literatura uma abordagem objetiva deste ponto. Não obstante, nós nos propomos a introduzir esta hipótese sob a assunção de que genes representem alguns dos mais importantes (senão o mais importante) substratos da determinação não estocástica da razão sexual. Em primeiro lugar, é de se ter em vista que grande parte das interações bioquímicas da via final são reguladas por genes; em segundo lugar, neoplasias associadas a alterações na razão sexual (Rune Jacobsen, et al., 2000) e envenenamentos por chumbo, boro e outros agentes tóxicos associados à diminuição da razão sexual, influenciam o ligamento/ desligamento de genes (James, 1997a, 1997b; Mocarelli, et al., 2000; Ryan, et al., 2002). Em terceiro, más formações do sistema nervoso se distribuem de maneira não estocástica entre os sexos (Lemire & Pendergrass, 2002), sugerindo a possibilidade de que mutações comuns atuem em ambos os níveis (sexual e nervoso).
2. Das vias intermediárias às vias moleculares implicadas na razão sexual DE nascidos humanos
Para dar conta da derradeira modelagem da via ecológico ? molecular faz-se imperiosa a circunscrição prévia da referida via final. Esta compreende dois recortes: em nível primário se refere às condições bioquímicas da determinação do percentual andro espermático, associado às condições bioquímicas do líquido folicular implicadas na suscetibilidade à fecundação por androspermas e gimnospermas. Em nível secundário, o fenômeno reduz-se às contingências maternas associadas à homeostase fetal.
Não obstante, e tal como previamente assinalado, é de se ter em vista a existência de variáveis intermediárias entre as ecológicas e as derradeiras vias finais moleculares, o que naturalmente converge à premissa de que existe um conjunto limitado de fenômenos moleculares não finalistas, candidatas a influenciar o derradeiro comportamento da via finalista. A importância dos mesmos é paradigmática: a via finalista é por natureza homeostática, de modo que toda alteração será por alterações em percentuais de elementos naturalmente presentes no mileu parental. Investigar os mais fortes candidatos é uma função da prospecção por vias intermediárias moleculares.
2.a Os principais dados que vinculam determinantes paternos à Razão sexual são:
i. Dados relativos a intoxicações/envenenamentos potencialmente capazes de diminuir a razão sexual através da diminuição de funções testiculares em associação com quedas nos níveis de testosterona (James, 1997a, 1997b; Mocarelli, et al., 2000; Ryan, et al., 2002).
ii. câncer testicular (neoplasia das células germinativas intratubulares) parece estar de algum modo envolvido diminuição a razão sexual (Jacobsen, et al., 2000). Não obstante, levantamos a necessidade de não se assumir dogmaticamente a perspectiva uma relação diretiva, posto ter sido demonstrado que irmãos não afetados de portadores de tais neoplasias apresentam alterações reprodutivas (declínio da progênie e diminuição da razão sexual em gêmeos monozigóticos), ao passo que suas irmãs permanecem reprodutivamente intactas (Richiardi & Akre, 2005). Nossa revisão de experimentos sugere que o fator mais determinante seja genético, criticamente representado por mutações no gene que sintetiza o receptor androgênico, entre as quais se destacam pares de tripletes CAG/GGC no exon 1 do gene TGCT do cromossomo XQ27, as quais diminuem a atividade transcripcional (?RNAm) necessária para a síntese do receptor. Mais uma vez, destacamos a importância desta consideração dada a inexistência de referências na literatura acerca da hipótese (que nos parece consistente) de que os genes de um indivíduo atuam na determinação da razão sexual.
iii. Antígenos HLA são glicoproteínas membramáticas que exercem a função leucocitária e que frequentemente se encontram em níveis aumentados em associação a doenças autoimunes. Particularmente a glicoproteína HLA-B27 parece estar relacionada à síndrome de Reiter, artrite reumatoide, psoríase, espondilite anquilosante e outras, assim como a taxas aumentadas de testosterona em homens. Em relação a estas, sabe-se que a glicoproteína HLA-B44 apresenta função protetiva, ao mesmo tempo em que também apresenta caráter antiandrogênico. Um estudo de 1991 sugeriu que portadores de artrite reumatoide gerem menores razões sexuais, assim como portadores de síndrome de Reiter gerem razões aumentadas (James, 1991), possivelmente em associação proximal com estes fatores leucocitários e em relação indireta para com o nível de testosterona do pai.
iv. Em suma, o nível de testosterona paterno é o principal candidato a representante masculino das alterações não estocástica da razão sexual entre nascidos humanos. Ao que se pode acrescentar o fato de que homens subférteis (baixa testosterona) produzem razões sexuais menores do que a média, efeito o qual se inverte quando estes são tratados com metil-testosterona (James, 1990), enquanto efeitos correlatos em modelos animais fazem-se bem conhecidos (Goerlich, Dijkstra, Schaafsma, & Groothuis, 2009; Grant, et. al, 2008; Zlabek, et. al, 2009).
De todo o modo, mais uma vez ressaltamos que a relação proximal pode exigir outros fatores, entre os quais a presença de comportamento alterado de genes, tal como sugeriu um estudo que revelou a presença de repetições diminuídas de microssatélites CAG no gene que codifica o receptor androgênico, em uma população (japonesa) de portadores de espondilite anquilosante. Isto é, pode não ser o caso de se fechar o veredito sobre a influência pontual dos níveis de testosterona teciduais, devendo-se manter em vista as peculiaridades da atividade genética e determinantes epigênicos (i.e. metilizações específicas) no âmbito da síntese e degradação de receptores, posto que tais frequentemente regulam múltiplos fatores, podendo alguns passar injustamente despercebidos sobre o destaque dado à testosterona.
Por fim, destacamos que diversos estudos apontam que as influências maternas são mais importantes do que as paternas, vicissitude a qual nos parece factível, tal como desenvolveremos na próxima seção (Coney & Mackey, 1998; Grant, 2007).
2.b Os principais dados que vinculam determinantes maternos à razão sexual são:
i. Doenças celíacas diminuem a razão sexual, ao alterarem o nível de vários hormônios, entre os quais se destaca a testosterona (James, 2001a).
ii. Anomalias placentárias: gravidez extrauterina e placenta com penetração extrema da decídua (placenta acreata) parecem estar ligadas à diminuição da razão sexual; desprendimento prematuro da placenta (placenta abrupta) e hipertensão gravídica (toxemia) parecem estar ligados ao aumento da razão sexual (James, 1995). Os mecanismos para tanto não são claros ainda.
iii. Circunferência da cintura parece estar, dentro de determinados parâmetros, diretamente relacionada ao aumento da razão sexual, possivelmente desde efeitos testosteronérgicos (Manning, Anderton, & Washington, 1996).
2.c Os principais dados que vinculam determinantes maternos e paternos à razão sexual são:
Estresse: o estresse pode ser definido como uma resposta involuntária a eventos ameaçadores e/ou ameaçadores, em diversos níveis (dobson & smith, 2000). neste sentido, trata-se de um conceito bastante amplo e pouco informativo, que pode substituir cada um dos vetores das alterações não estocásticas da razão sexual, em representações estruturais frouxas (estresse químico, mecânico, psicológico, etc.).
Assim é que, evitando o reducionismo de um aglutinador geral das alterações pontuais da razão sexual, limitamo-nos a utilizar o conceito de estresse em sentido psicológico, do que desponta a máxima de que tal seja a força motriz por trás de alterações da razão sexual associadas a guerras e catástrofes naturais, dada a conhecida associação entre este e alterações significativas da homeostase, com aumento da atividade simpática, entre outros efeitos (Hjemdahl, Freyschuss, Juhlin-Dannfelt, & Linde, 1983).
Os efeitos orgânicos da representação mental duradoura dos eventos críticos atingem a síntese de esteroides, onde então se configura o mais significativo elo intermediário desta cascata de eventos inexoravelmente somático-mental. Neste âmbito, a importância do cortisol é elementar, sob o mote da relação geral entre os glicocorticoides do eixo HPA e os esteroides sexuais do eixo HPG (Viau, 2002). De maneira mais específica, fazem-se proeminentes a cascata bioquímica iniciada na glândula pituitária, onde o cortisol inibe as respostas das células gonadotróficas e assim leva a uma diminuição direta da capacidade reprodutiva por via da inibição da gonadotrofina coriônica e do hormônio luteinizante (Tilbrook, Turner, & Clarke, 2000), e a cascata da ação bidimensional da testosterona sobre a síntese da vasopressina/síntese da corticotrofina e então do próprio nível de cortisol (Popma, et al., 2007), convergindo para a conformação de um sistema de retroalimentação bioquímica.
3. Vias finais da determinação não estocástica da razão sexual entre nascidos humanos
Tal como assinalado, a via final é representada por interações hormonais que em última análise estabelecerão a probabilidade de que o óvulo seja fertilizado por um androsperma, mais a probabilidade de aborto espontâneo vezes a razão sexual dos abortos, divido pela razão sexual dos abortos da média da população.
Segundo James, a representação das variáveis não estocásticas da razão sexual humana em termos hormonais pode ser dada pela equação (T + E)/(G + P), onde T: testosterona, E: estrógeno, G: gonadotrofina coriônica, e P: progesterona (James, 1997c).
Não obstante, é de se ter em vista que, ao que tudo indica, a influência masculina limita-se a uma eventual alteração na razão androsperma/ gimnosperma, talvez em função de ser o zigoto androespermático mais suscetível ao estresse químico. não há, por exemplo, nenhum registro na literatura de que o padrão enzimático (hidrolítico) dos acromossomos possa estar de alguma maneira relacionadas à presença de cromossomo x ou y no cariótipo do espermatozoide, que assim poderia atuar seletivamente sobre a fecundação, desde uma igualmente hipotética vantagem para o vencimento da coroa radiata e da zona pelúcida do óvulo ao longo da via final – hipótese a qual, eventualmente, poderá despontar como forte candidata para uma eventual participação masculina neste âmbito e que sugerimos como futuro manancial de estudos e eventuais descobertas.
Considerando assim que o fenômeno em questão ganha forma na região ampular da trompa de falópio, por força da exposição do óvulo a espermatozoides e que o cânone é o recorte na suscetibilidade do óvulo ao vencimento da coroa radiata, faz-se importante considerar que os fatores implicados na via final sejam exclusivamente os hormônios femininos representados em tal conjuntura do ambiente folicular.
Neste sentido os dois hormônios mais significativos para as alterações não estocásticas da razão sexual parecem ser: gonadotrofina coriônica (HCG) e a testosterona materna. O estrógeno é relacionado à razão sexual em outras espécies, tais como peixes (Hill & Janz, 2003), tartarugas (Bergeron, Willingham, & Osborn, 1999) e cachorros selvagens (creel, creel, & Monfort, 1998), mas faltam dados sobre humanos, ainda que existam especulações a respeito (James, 1990), na mesma linha em que a equação de James previamente apresentada representa uma especulação à espera de um mínimo de base experimental.
Índices aumentados de hcG são frequentemente encontrados em reproduções induzidas, condição na qual a maior parte da literatura sugere que nasçam mais meninas. A testosterona se encontra frequentemente aumentada em mulheres sob estresse crônico (Grant & irwin, 2005), as quais tendem a mostrar razão sexual aumentada, havendo mesmo quem sugira ser a testosterona materna o único índice hormonal importante para as alterações não estocásticas da razão sexual (Grant, 2007).
de fato, a despeito do fato de que uma análise abrangente da literatura sugira que níveis elevados de gonadotrofina coriônica igualmente afetem a razão sexual (James, 1980, 1990; Santolaya-Forgas, Meyer, Burton, & Scommegna, 1997), neste caso a diminuindo (Grant, 1998), os dados são muito mais consistentes no que tange àquela -não sendo desprezível o fato de que o único experimento que testou aumentar os níveis de HCG durante fertilização in vitro chegou à conclusão que de que isto não muda a razão sexual (Zhang, et. al, 2008).
Um último indicativo experimental do papel proeminente da testosterona faz-se oriundo da etologia, conforme alguns estudos apontam que a razão sexual se relaciona com a posição da mãe no ranking social em primatas e então ao nível de testosterona circulante (Blanchard, Festa-Bianchet, Gaillard, & Jorgenson, 2005; Maestripieri & Blake, 2002; Nevison, 1997; Schino, 2004), padrão o qual parece se reiterar entre humanas (Grant, 1996).
Dito isto, destacamos o caráter controverso desta hipótese e de achados pitorescos associados, tal como o que aponta que mulheres que exercem profissões mais masculinizadas (i.e. engenharia) têm maior proporção de filhos homens do que as que exercem profissões menos masculinizadas
(i.e. enfermagem) (Kanazawa & Vandermassen, 2005), em função de regulação testosteronérgica, perspectiva a qual o estatístico Andrew Gelman ressalta como matematicamente insustentável (Gelman, 2007).
Por fim, não deixa de ser digno de nota que uma busca no PubMed para testosterona e razão sexual retorna 1433 referências, enquanto gonadotrofina coriônica e razão sexual retorna 67 referências (gonadotrofina e razão sexual retorna 577); razão sexual em primatas e nível hormonal e testosterona retorna 167 referências, enquanto razão sexual em primatas e nível hormonal e gonadotrofina retorna apenas 64.
Por força destas vicissitudes, procederemos focados na dinâmica da flutuação testosteronérgica folicular como via final das variações não estocásticas da razão sexual, sob a máxima de que o futuro pode trazer reviravoltas a esta perspectiva.
Para os nossos propósitos, o nível testosteronérgico folicular pode ser representado como a saída de um sistema onde interagem dois conjuntos de variáveis: fatores que aumentam o seu nível e fatores que o diminuem, tais podendo ser alinhavados sob o conceito genérico e (propositalmente) pouco informativo de estressores.
Denominemos a saída quando da ação exclusiva do primeiro conjunto como x e a saída quando da ação exclusiva do segundo como x; estruturalmente este problema pode ser representado a partir de um só conjunto (testosterona/estresse) x, sendo x definido como fenômeno que ocorre quando da não realização da condição x. Isto é, utilizamos a tautologia de que o aumento do nível de testosterona é justamente aquilo que esperamos que se realize quando não ocorrer a diminuição da mesma e mesmo assim atuar um mecanismo de determinação não estocástica da razão sexual entre nascidos para proceder com uma representação molecular da via final, em que apenas fatores contribuem para uma das saídas (x) são representados.
Isto por sua vez dá origem à cascata bioquímica abaixo, a qual foi implementada utilizando módulo search pathway do KEGG (http://www.genome.jp/kegg/tool/search_pathway.html):
Para se abordar a cadeia proposta de maneira significativa, é de se começar pela consideração de que tanto o principal hormônio relacionado ao estresse (cortisol) quanto a testosterona são esteroides e os poucos estudos que avaliaram a relação de ambos in situ, concluíram por relações diretas (Sarkar, 2008; Sarkar, et. al, 2007), tal nos parecendo ser a vicissitude mais significativa de todas.
A relação entre testosterona folicular e estresse se faz mediada pela liberação de glicocorticoides (desde a ação não representada de núcleos hipotalâmicos e pituitários), a qual converge à síntese de corticoesterona (na zona glomerular do córtex adrenal) e cortisol (tanto na região fasciculada quanto na glomerulada da mesma área).
A corticoesterona se converterá em aldotestosterona, que é um hormônio importante na reabsorção hepática, o qual possivelmente tem como principal via ao comportamento, a estimulação da produção de vasopressina (associada ao acasalamento) e o cortisol ativará diversos sistemas de ataque e fuga (gástrico, respiratório, valência/saliência mental), através da via que passa pela síntese cortisona. Sabe-se que a cortisona inibe o desenvolvimento de doenças autoimunes, as quais parecem estar relacionadas a alterações da razão sexual, como previamente mencionado.
Em nível bastante elementar o gene cyP11b2 (18-hidroxilase) sintetiza proteínas enzimáticas da família P450, as quais estão entre as principais responsáveis pela síntese dos esteroides da cadeia da aldoesterona.
No ramo oposto, o principal gene ao papel de agente na determinação da razão sexual humana é o HSD11B1 (11-hidroxiesteroide dehidrogenase) responsável pela redução enzimática do cortisol desde a cortisona, através da cadeia representada abaixo do mesmo (em amarelo), atingindo a síntese do metabólito cortolone e o glucorinídeo (associado à excreção tóxica), que foram associados ao nível de testosterona e cortisol em humanos à luz de possível regulação gênica (Pritchard, et al., 1998). Destacamos a inexistência de registros na literatura acerca da associação entre polimorfismos no HSD11B1 e alterações da razão sexual e o caráter potencialmente de tais estudos (Ref Seq/Entrez).
4. Testosterona e razão sexual
A popularidade da associação entre testosterona e razão sexual em ao menos alguma espécie pode ser medida pelo fato de existirem 1435 publicações indexadas no PubMed sob ambos os conceitos (and).
Como não poderia ser diferente, tal número de publicações se traduz não apenas como possibilidade de muitos insights, oriundos de associações oportunas de ideias e achados, como impossibilidade de se fornecer uma sistematização crítica dos principais focos destas pesquisas.
Para solucionar esta questão, apresentamos abaixo um mapa temático do campo testosterona e razão sexual, construído através do software Omniviz, módulo Thememap. O princípio básico para a divisão temática (cuja importância relativa no campo está relacionada ao volume ocupado no mapa) é a aplicação de algoritmos para a determinação de padrões recorrentes ao longo dos metadados, assim propiciando o agregamento temático das várias perspectivas (clusters), em associação a tabelas de conceitos para cada unidade temática; nesta organização. Em outras palavras: primeiro o software define os conceitos essenciais e depois constrói um mapa desde o valor estatístico e as relações entre eles. Neste, volume significa quantidade de publicações, enquanto verticalidade significa concentração temporal e autoral (i.e. um assunto pode ser muito explorado, porém ter uma representação dispersa conquanto seja antigo e frequentemente revisitado por autores diversos).
Tal como sugerido pelo mapa, as principais associações conceituais envolvem a concepção de que o nível de exposição materna aos esteroides determina a razão sexual, o que concluímos dada o destaque das associações steriod/ratio/female/expose/concentration, presente no maior cluster;male/ratio/female/hormone/concentration/offspring, presente no segundo maior. Paralelamente, aponta-se para a existência de estudos avaliando os efeitos da exposição de fetos de animais aos hormônios maternos (male/female/fetus/litter/exposure/offspring/ratio) e outros que apontam para a importância da pesquisa com animais, com destaque para ratos e pássaros.
Conclusão
Neste estudo revisamos sistematicamente alguns dos principais aspectos relacionados à determinação da razão sexual, em seus aspectos demográficos e fisiológicos e, finalmente, apresentamos um mapa conceitual do campo, baseado no uso de um avançado sistema de text mining (mineração da literatura).
As principais conclusões do estudo são que a razão sexual possui determinação fisiológica, envolvendo diversas cascatas hormonais, com ênfase para aquelas envolvendo a regulação do nível de testosterona materno, no líquido folicular. Esta regulação sofre efeitos top-down do cortisol e outros derivados desta via metabólica, os quais podem ser liberados em função de estresse psicológico, explicando assim as alterações na razão sexual em situações de calamidades públicas e guerras. Neste sentido, é fundamental ter em vista o papel das interações entre cortisol e testosterona, conforme delineado na Figura 2 e na discussão subsequente.
Este é um campo muito pouco explorado, sobretudo em se considerando a imensa importância e generalidade de suas questões para toda a espécie humana.
Dito isto, é importante estarmos atentos às limitações do estudo: as vias moleculares que exploramos foram baseadas em dados do KEEG, que não conhecemos em detalhes, enquanto o mapa de estudos indexados no Pubmed não possui contornos estatísticos precisos, antes servindo para uma visão geral dos subtemas que cruzam razão sexual e testosterona.
Referências
Abu-Musa, A. A., Usta, I. M., Yunis, K., & Nassar, A. H. (2009). Effect of 1-month war in Lebanon on sex ratio. Fertility and Sterility, 91(4), 1571-1573. [ Links ]
Allan, B. B., Brant, R., Seidel, J. E., & Jarrell, J. F. (1997). Declining sex ratios in Canada. CMAJ, 156(1), 37-41. [ Links ]
Ansari-Lari, M., & Saadat, M. (2002). Changing sex ratio in Iran, 1976-2000. J Epidemiol Community Health, 56(8), 622-623. [ Links ]
Astolfi, P., & Zonta, L. A. (1999). Reduced male births in major Italian cities. Hum. Reprod., 14(12), 3116-3119. [ Links ]
Bergeron, J., Willingham, E., & Osborn, C. (1999). Developmental synergism of steroidal estrogens in sex determination. Environmental Health Perspectives, 107(2), 93. [ Links ]
Blanchard, P., Festa-Bianchet, M., Gaillard, J.-M., & Jorgenson, J. T. (2005). Maternal condition and offspring sex ratio in polygynous ungulates: a case study of bighorn sheep. Behav. Ecol., 16(1), 274-279. [ Links ]
Cagnacci, A., Renzi, A., Arangino, S., Alessandrini, C., & Volpe, A. (2004). Influences of maternal weight on the secondary sex ratio of human offspring. Hum. Reprod., 19(2), 442-444. [ Links ]
Coney, N. S., & Mackey, W. C. (1998). The woman as final arbiter: a case for the facultative character of the human sex ratio. Journal of Sex Research, 35, 169-175. [ Links ]
Creel, S., Creel, N., & Monfort, S. (1998). Birth order, estrogens and sex-ratio adaptation in . Animal reproduction science, 53, 315-320. [ Links ]
Di Renzo, G. C., Rosati, A., Sarti, R. D., Cruciani, L., & Cutuli, A. M. (2007). Does fetal sex affect pregnancy outcome? Gender Medicine, 4(1), 19-30. [ Links ]
Dobson, H., & Smith, R. (2000). What is stress, and how does it affect reproduction? Animal reproduction science, 60, 743-752. [ Links ]
Fukuda, M., Fukuda, K., Shimizu, T., & Moller, H. (1998). Decline in sex ratio at birth after Kobe earthquake. Human Reproduction, 13(8), 2321-2322. [ Links ]
Gelman, A. (2007). Letter to the editors regarding some papers of Dr. Satoshi Kanazawa. Journal of Theoretical Biology, 245(3), 597-599. [ Links ]
Goerlich, V., Dijkstra, C., Schaafsma, S. M., & Groothuis, T. G. (2009). Testosterone has a long term effect on primary sex ratio of first eggs in pigeons -- in search of a mechanism. Gen Comp Endocrinol. [ Links ]
Graffelman, J., & Hoekstra, R. F. (2000). A statistical analysis of the effect of warfare on the human secondary sex ratio. Human Biology, 72(3), 433-445. [ Links ]
Grant, V. (1996). Sex determination and the maternal dominance hypothesis. ESH RE, 11, 2371-2375. [ Links ]
Grant, V. (1998). Maternal personality, evolution and the sex ratio: do mothers control the sex of the offspring: London, UK: Routledge. [ Links ]
Grant, V. J. (2007). Could maternal testosterone levels govern mammalian sex ratio deviations? Journal of Theoretical Biology, 246(4), 708-719. [ Links ]
Grant, V. J., & Irwin, R. J. (2005). Follicular fluid steroid levels and subsequent sex of bovine embryos. Journal of Experimental Zoology Part A: Comparative Experimental Biology, 303(12), 1120-1125. [ Links ]
Grant, V. J., Irwin, R. J., Standley, N. T., Shelling, A. N., & Chamley, L. W. (2008). Sex of bovine embryos may be related to mothers preovulatory follicular testosterone. Biol Reprod, 78(5), 812-815. [ Links ]
Gupta, M., & Bhat, P. (1997). Fertility decline and increased manifestation of sex bias in India. Population Studies, 51(3), 307-315. [ Links ]
Hill, R., & Janz, D. (2003). Developmental estrogenic exposure in zebrafish (Danio rerio): I. Effects on sex ratio and breeding success. Aquatic toxicology, 63(4), 417-429. [ Links ]
Hjemdahl, P., Freyschuss, U., Juhlin-Dannfelt, A., & Linde, B. (1983). Differentiated sympathetic activation during mental stress evoked by the Stroop test. Acta physiologica Scandinavica. Supplementum,
527, 25-29.
Jacobsen, R., Bostofte, E., Engholm, G., Hansen, J., Skakkebak, N. E., & Moller, H. (2000). Fertility and offspring sex ratio of men who develop testicular cancer: a record linkage study. Hum. Reprod., 15(9), 1958-1961. [ Links ]
Jacobsen, R., Moller, H., & Mouritsen, A. (1999). Natural variation in the human sex ratio. Hum. Reprod., 14(12), 3120-3125. [ Links ]
James, W. H. (1980). Gonadotrophin and the human secondary sex ratio. Br Med J, 281(6242), 711-712. [ Links ]
James, W. H. (1984). Seasonality in the sex ratio of US Black births. Annals of Human Biology, 11(1), 67 - 69. [ Links ]
James, W. H. (1987). The human sex-ratio Human Biology, 59(5), 721-752. [ Links ]
James, W. H. (1990). The hypothesized hormonal control of human sex ratio at birth--an update. J Theor Biol, 143(4), 555-564. [ Links ]
James, W. H. (1991). Sex ratios and hormones in HL A related rheumatic diseases. Ann Rheum Dis, 50(6), 401-404. [ Links ]
James, W. H. (1995). Sex ratios of offspring and the causes of placental pathology. Hum. Reprod., 10(6), 1403-1406. [ Links ]
James, W. H. (1996). Interpregnancy intervals, high maternal age and seasonal effects on the human sex ratio. Hum Reprod, 11(1), 7-8. [ Links ]
James, W. H. (1997a). Parental HL A genes, hormone levels and offspring sex ratios. Hum Hered, 47(1), 60. [ Links ]
James, W. H. (1997b). Paternal lead exposure, offspring birth weight, and sex ratio. Am J Ind Med, 32(3), 315-316. [ Links ]
James, W. H. (1997c). A potential mechanism for sex ratio variation in mammals. Journal of Theoretical Biology, 189(3), 253-255. [ Links ]
James, W. H. (2001a). Hormones and offspring sex ratios associated with celiac disease. The American Journal of Gastroenterology, 96(7), 2266-2267. [ Links ]
James, W. H. (2001b). Variation in the human sex ratio at birth with maternal age, parity and placental pathology. Hum. Reprod., 16(10), 2246-2247. [ Links ]
Jha, P., Kumar, R., Vasa, P., Dhingra, N., Thiruchelvam, D., & Moineddin, R. (2006). Low male-to-female sex ratio of children born in India: national survey of 1 1 million households. The Lancet, 367(9506), 211-218. [ Links ]
Johansson, S., & Nygren, O. (1991). The missing girls of China: A new demographic account. The Population and Development Review, 35-51. [ Links ]
Joseph, K., Wilkins, R., Dodds, L., Allen, V., Ohlsson, A., Marcoux, S., et al. (2005). Customized birth weight for gestational age standards: Perinatal mortality patterns are consistent with separate standards for males and females but not for blacks and whites. BMC Pregnancy and Childbirth, 5(1), 3. [ Links ]
Juntunen, K., Kvist, A., & Kauppila, A. (1997). A shift from a male to a female majority in newborns with the increasing age of grand grand multiparous women. Hum. Reprod., 12(10), 2321-2323. [ Links ]
Kanazawa, S., & Vandermassen, G. (2005). Engineers have more sons, nurses have more daughters: an evolutionary psychological extension of Baron-Cohens extreme male brain theory of autism. Journal of Theoretical Biology, 233(4), 589-599. [ Links ]
Kraemer, S. (2000). The fragile male. BMJ: British Medical Journal, 321(7276), 1609-12. [ Links ]
Lichtenfels, A. J. d. F. C. (2006). Associação entre os níveis ambientais de poluição atmosférica e mudanças na razão sexual na cidade de São Paulo: uma abordagem epidemiológica e experimental. Universidade de São Paulo. Faculdade de Medicina. [ Links ]
Macmahon, B., & Pugh, T. F. (1954). Sex ratio of white births in the United States during the Second World War. Am J Hum Genet, 6(2), 284-292. [ Links ]
Maestripieri, D., & Blake, B. H. (2002). Maternal dominance rank and age affect offspring sex ratio in pigtail macaques. Journal of Mammalogy, 83(2), 563-568. [ Links ]
Manning, J. T., Anderton, R., & Washington, S. M. (1996). Womens waists and the sex ratio of their progeny: evolutionary aspects of the ideal female body shape. Journal of Human Evolution, 31(1), 41-47. [ Links ]
Melnikov, V. N., & Grech, V. (2003). Seasonality of live birth sex ratio in south western Siberia, Russia, 1959-2001. J Epidemiol Community Health, 57(6), 471-472. [ Links ]
Miller, B. (1997). The endangered sex: Neglect of female children in rural North India. Oxford: Oxford University Press. [ Links ]
Miura, T., Nonaka, K., Shimura, M., & Nakamura, I. (1983). A study of the sex ratio of first-born According to the mothers month of birth. Archives of Gynecology and Obstetrics, 233(4), 263-266. [ Links ]
Mocarelli, P., Brambilla, P., Gerthoux, P. M., Jr, D. G. P., & Needham, L. L. (1996). Change in sex ratio with exposure to dioxin. The Lancet, 348(9024), 409-409. [ Links ]
Mocarelli, P., Gerthoux, P., Ferrari, E., Patterson, D., Kieszak, S., Brambilla, P., et al. (2000). Paternal concentrations of dioxin and sex ratio of offspring. The Lancet, 355(9218), 1858-1863. [ Links ]
Mocarelli, P., Gerthoux, P. M., Ferrari, E., Patterson, J. D. G., Kieszak, S. M., Brambilla, P., et al. (2000). Paternal concentrations of dioxin and sex ratio of offspring. The Lancet, 355(9218), 1858-1863. [ Links ]
Moller, H. (1996). Change in male:female ratio among newborn infants in Denmark. Lancet, 348(9030), 828-829. [ Links ]
Møller, H. (1996). Change in male:female ratio among newborn infants in Denmark. The Lancet, 348(9030), 828-829.
Mori, I., Ogawa, Y., Koda, S., Kumagai, S., & Ueno, M. (2004). the Sex Ratio in the Offspring of Municipal Solid Waste Incinerator Workers. Epidemiology, 15(4), S118-S119. [ Links ]
Nations, U. (2009). Human Development Report 2007/2008. Genebre: United Nations Development Programme [ Links ]
Navara, K. J. (2009). Humans at tropical latitudes produce more females. Biology Letters, [ Links ] -.
Nevison, C. (1997). Social rank and birth sex ratios in rhesus monkeys (Macaca mulatta). Applied Animal Behaviour Science, 51(3-4), 285-291. [ Links ]
P. Sarkar, K. B., T. G. OConnor, V. Glover,. (2008). Maternal Antenatal Anxiety and Amniotic Fluid Cortisol and Testosterone: Possible Implications for Foetal Programming. Journal of Neuroendocrinology, 20(4), 489-496. [ Links ]
Parazzini, F., La Vecchia, C., Levi, F., & Franceschi, S. (1998). Trends in male:female ratio among newborn infants in 29 countries from five continents. Hum. Reprod., 13(5), 1394-1396. [ Links ]
Park, C., & Cho, N. (1995). Consequences of son preference in a low-fertility society: imbalance of the sex ratio at birth in Korea. Population and Development Review, 59-84. [ Links ]
Polasek, O., Kolcic, I., Kolaric, B., & Rudan, I. (2005). Sex ratio at birth and war in Croatia (1991-1995). Hum Reprod, 20(9), 2489-2491. [ Links ]
Popma, A., Vermeiren, R., Geluk, C. A. M. L., Rinne, T., van den Brink, W., Knol, D. L., et al. (2007). Cortisol Moderates the Relationship between Testosterone and Aggression in Delinquent Male Adolescents. Biological Psychiatry, 61(3), 405-411. [ Links ]
Pritchard, J., Despres, J. P., Gagnon, J., Tchernof, A., Nadeau, A., Tremblay, A., et al. (1998). Plasma adrenal, gonadal, and conjugated steroids before and after long-term overfeeding in identical twins.J Clin Endocrinol Metab, 83(9), 3277-3284. [ Links ]
Richiardi, L., & Akre, O. (2005). Fertility Among Brothers of Patients with Testicular Cancer. Cancer Epidemiol Biomarkers Prev, 14(11), 2557-2562. [ Links ]
Ryan, J., Amirova, Z., & Carrier, G. (2002). Sex ratios of children of Russian pesticide producers exposed to dioxin. Environmental Health Perspectives, 110(11), A699. [ Links ]
Saadat, M. (2008). Decline in sex ratio at birth after bam (kerman province, southern iran) earthquake. Journal of Biosocial Science, 40(06), 935-937. [ Links ]
Santolaya-Forgas, J., Meyer, W. J., Burton, B. K., & Scommegna, A. (1997). Altered newborn gender distribution in patients with low mid-trimester maternal serum human chorionic gonadotropin (MShCG). J Matern Fetal Med, 6(2), 111-114. [ Links ]
Sarkar, P., Bergman, K., Fisk, N. M., OConnor, T. G., & Glover, V. (2007). Amniotic fluid testosterone: relationship with cortisol and gestational age. Clin Endocrinol, 67(5), 743-747. [ Links ]
Schino, G. (2004). Birth sex ratio and social rank: consistency and variability within and between primate groups. Behav. Ecol., 15(5), 850-856. [ Links ]
Tilbrook, A., Turner, A., & Clarke, I. (2000). Effects of stress on reproduction in non-rodent mammals: the role of glucocorticoids and sex differences. Rev Reprod, 5(2), 105-113. [ Links ]
Tuljapurkar, S., Li, N., & Feldman, M. (1995). High sex ratios in Chinas future. Science, 267(5199), 874-876. [ Links ]
Ulizzi, L. A. Z. (1993). Sex ratio and natural selection in humans: a comparative analysis of two Caucasian populations. Annals of Human Genetics, 57(3), 211-219. [ Links ]
Lemire, R. J., & Pendergrass, T. W. (2002). Sex Ratios in Congenital Malformations of the Central Nervous System. Pediatric Neurosurgery, 36(1), 2-7. [ Links ]
Viau, V. (2002). Functional Cross-Talk Between the Hypothalamic-Pituitary-Gonadal and -Adrenal Axes. Journal of Neuroendocrinology, 14(6), 506-513. [ Links ]
van der Pal-de Bruin, K. M., Verloove-Vanhorick, S. P., & Roeleveld, N. (1997). Change in male:female ratio among newborn babies in Netherlands. Lancet, 349(9044), 62. [ Links ]
Wilson, E. O. (1978). What is sociobiology? Society, 15(6), 10-14. [ Links ]
Zhang, L., Du, W., Lin, X., Zhang, A., & Chen, H. (2008). Progesterone and 17beta-estradiol, but not follicle stimulating hormone, alter the sex ratio of murine embryos fertilized in vitro. Theriogenology, 69(8), 961-966. [ Links ]
Zlabek, V., Randak, T., Kolarova, J., Svobodova, Z., & Kroupova, H. (2009). Sex differentiation and vitellogenin and 11-ketotestosterone levels in chub, Leuciscus cephalus L., exposed to 17 betaestradiol and testosterone during early development. Bull Environ Contam Toxicol, 82(3), 280-284. [ Links ]
Álvaro Machado Dias
Laboratório de Neuroimagem LI M-21, Instituto de Psiquiatria da Universidade de São Paulo, R. Dr . Ovídio Pires de Camp os, 785, Caixa Postal 3671, CEP 01060-970 - São Paulo – SP, Brasil. E-mail: alvaromd@usp.br
Data de recepção / reception date: 08/07/2013
Data de aprovação / approval date: 08/01/2014

