










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.42 no.2 Lisboa jun. 2019
https://doi.org/10.19084/rca.15433
ARTIGO
Biomassa e atividade microbiana em solo cultivado com milho consorciado com leguminosas de cobertura
Biomass and microbial activity in soil cultivated with maize intercropped with soil cover legumes
Tais dos S. Araujo1, Anderson de S. Gallo2,*, Faguiner dos S. Araujo3, Lilian C. dos Santos4, Nathalia de F. Guimarães 2 e Rogério F. da Silva5
1 Universidade Estadual de Mato Grosso do Sul, Campo Grande, Mato Grosso do Sul, Brasil
2 Departmento de Solos, Instituto de Agronomia, Universidade Federal Rural do Rio de Janeiro, Seropédica, Rio de Janeiro, Brasil
3 Universidade Estadual de Mato Grosso do Sul, Medianeira, Paraná, Brasil
4 Universidade Estadual de Mato Grosso do Sul, Santa Tereza do Oeste, Paraná, Brasil
5 Universidade Estadual de Mato Grosso do Sul, Glória de Dourados, Mato Grosso do Sul, Brasil
(*E-mail: andersondsgallo@hotmail.com)
RESUMO
O carbono orgânico do solo e a atividade da biomassa microbiana têm sido utilizados como indicadores de qualidade do solo em agroecossistemas, visto que, por serem sensíveis, são capazes de refletir as alterações causadas pelos diferentes sistemas de manejo do solo. Objetivou-se neste trabalho avaliar as alterações na biomassa microbiana e sua atividade em solo sob cultivo de milho consorciado com leguminosas. O delineamento experimental adotado foi blocos casualizados, em esquema fatorial (6 x 2), com quatro repetições. Os tratamentos constituíram de arranjos de milho consorciado com leguminosas: monocultivo de milho; milho consorciado com feijão-de-porco; com crotalária; com mucuna-preta; guandu-anão e com feijão-caupi, associados a duas profundidades de amostragem (0,0-0,10 m e 0,10-0,20 m). As variáveis avaliadas foram: carbono da biomassa microbiana (C-BMS), respiração basal (C-CO2), quociente metabólico (qCO2), quociente microbiano (qMIC) e matéria orgânica do solo (MOS). Na camada superficial, as coberturas de solo influenciaram todas as variáveis analisadas. Os consórcios de milho com crotalária e guandu-anão proporcionaram um carbono da biomassa microbiana e quociente microbiano semelhante à do solo na condição natural. O sistema com milho em monocultivo apresentou alto quociente metabólico, pressupondo condições de estresse. O C da biomassa microbiana do solo (C-BMS) apresenta maior qualidade na profundidade de 0,0-10 cm.
Palavras-chave: consórcio de culturas, indicadores bioquímicos, matéria orgânica do solo, quociente microbiano
ABSTRACT
Soil organic carbon and microbial biomass activity have been used as indicators of soil quality in agroecosystems, since they are sensitive to reflect changes caused by different soil management systems. This study evaluated changes in microbial biomass and its activity in soil under cultivation of maize intercropped with legumes. The experimental design was randomized blocks, factorial arrangement (6 x 2), with four replications. Treatments consisted of arrangements of maize intercropped with legumes: maize monoculture; maize intercropped with jack bean; with crotalaria; with velvet bean; pigeon pea and cowpea, associated to two sampling depths (0.0-0.10 m and 0.10-0.20 m). Variables evaluated included: microbial biomass carbon (C-BMS), basal respiration (C-CO2), metabolic quotient (qCO2), microbial quotient (qMIC) and soil organic matter (MOS). In the superficial layer, the soil cover influenced all analyzed variables. Corn intercropped with crotalaria and pigeon pea provided microbial biomass carbon and microbial quotient similar to soil in the natural condition. The system with monoculture maize showed high metabolic quotient, assuming stress conditions. The carbon of soil microbial biomass (C-BMS) shows better quality in the depth of 0.0-10 cm.
Keywords: intercropping; biochemical indicators; soil organic matter; microbial quotient
INTRODUÇÃO
O manejo inadequado do solo por meio das atividades agropecuárias pode com o tempo trazer graves consequências, exaurindo suas reservas orgânicas e minerais, transformando solos com grande potencial de produção em solos de baixa fertilidade (Duarte et al., 2014). Por outro lado, os sistemas integrados de produção agrícola, assim como outras práticas de manejo consideradas conservacionistas, apresentam-se como alternativas viáveis para a sustentabilidade do uso do solo (Silva et al., 2011). Uma das práticas conservacionistas capazes de promover a sustentabilidade dos solos agrícolas é a adubação verde, que consiste no cultivo e corte de plantas em plena floração, com ou sem incorporação da fitomassa, com a finalidade de aumento, preservação e/ou restauração da fertilidade do solo e da produtividade das culturas (Wutke et al., 2014; Brito et al., 2017).
A utilização de plantas de cobertura de solo e adubação verde é uma alternativa ecológica e econômica de manejar adequadamente o solo, possibilitando o equilíbrio das propriedades físicas, químicas e biológicas, que giram em torno do sistema solo-planta (Souza et al., 2008). A adubação verde, seja em consórcio ou sucessão com outras espécies, pode gerar quantidades de matéria seca suficientes para manter o solo coberto, aumentar o teor de matéria orgânica, contribuir na ciclagem de nutrientes, no armazenamento da água, na manutenção de temperatura mais baixa na camada superficial do solo e diminuir a evapotranspiração (Giongo et al., 2011). Além disso, favorece a ocorrência de organismos da fauna edáfica e a atividade microbiana, o que promove a ciclagem do nitrogênio e do carbono no solo (Almeida et al., 2016).
Recentemente tem crescido na comunidade científica o interesse por indicadores do funcionamento do sistema solo, baseados na atividade microbiana (Aragão et al., 2012). A biomassa microbiana é a principal responsável pela decomposição dos resíduos orgânicos, pela ciclagem de nutrientes e pelo fluxo de energia no solo, podendo ter influência tanto na transformação da matéria orgânica, quanto na estocagem do carbono e nutrientes minerais (Jenkinson e Ladd, 1981). Além disso, vem sendo utilizada como indicador de alterações e de qualidade do solo (Six et al., 2006), uma vez que esta associada às funções ecológicas do ambiente e são capazes de refletir rapidamente as mudanças de uso do solo (Jackson et al., 2003), pois qualquer estresse no sistema afeta a densidade, a diversidade e a atividade da microbiota (Matoso et al., 2012). Portanto, a avaliação da biomassa microbiana pode ajudar a orientar os produtores a manejarem seus solos de forma mais produtiva e sustentável (Aragão et al., 2012).
O conhecimento do impacto de sistemas conservacionistas de manejo sobre a biomassa microbiana do solo e sua atividade é importante. Essa informação pode contribuir para o estabelecimento de uma relação mais confiável entre o uso do solo e a sustentabilidade (Alves et al., 2011). No entanto, pouco se sabe sobre propriedades microbiológicas do solo em sistemas de consórcio de culturas (Song et al., 2007). Assim, o trabalho teve como objetivo, avaliar as alterações na biomassa microbiana e sua atividade em solo sob cultivo de milho consorciado com leguminosas.
MATERIAL E MÉTODOS
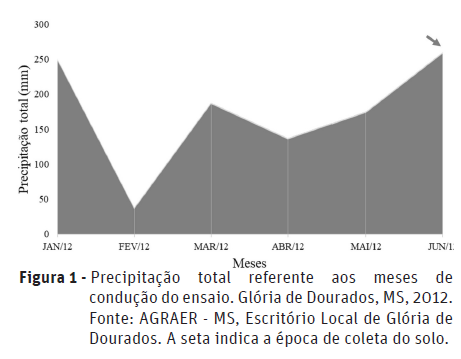
O trabalho foi realizado no campo experimental da Universidade Estadual de Mato Grosso do Sul (UEMS), município de Glória de Dourados, MS (22o22‘S e 54o30‘W, 400 m de altitude), num solo classificado como Argissolo Vermelho, de textura arenosa, com as seguintes características químicas na profundidade de 0 - 0,20 m: pH (H2O) = 5,5; P = 2,40 mg dcm-3; K = 0,19 cmolc dm-3; Ca = 0,8 cmolc dm-3; Mg = 0,4 cmolc dm-3; Al = 0,1 cmolc dm-3; H + Al: 2,9 cmolc dm-3 e matéria orgânica = 9,5 g kg-1. O clima de ocorrência da região, segundo a classificação de Köppen, é do tipo Aw, com estação quente e chuvosa no verão e moderadamente seca no inverno. A média de precipitação anual varia entre 1.400 e 1.700 mm. A temperatura média nos meses mais frios encontra-se em torno de 18°C e nos meses mais quentes fica em torno de 28°C, onde as temperaturas médias extremas atingem 35°C (Mercante et al., 2007). Os dados de precipitação pluvial total e média (mm) durante o período de estudo, encontram-se na Figura 1.
O delineamento experimental adotado foi blocos casualizados, em esquema fatorial (6 x 2), com quatro repetições. Os tratamentos constaram dos seguintes arranjos: monocultivo de milho (MS); milho consorciado com feijão-de-porco (Canavalia ensiformis (L.) D.C.) (M+FP); com crotalária (Crotalaria juncea L.) (M+CJ); com mucuna-preta (Mucuna aterrima (Piper & Tracy) Holland) (M+MP); com guandu-anão (Cajanus cajan (L.) Millsp) (M+GA) e com feijão-caupi (Vigna unguiculata (L.) Walp) (M+FC), associados a duas profundidades de amostragem do solo (0,0-0,10 m e 0,10-0,20 m). Foi incluída na avaliação uma área com fragmento de vegetação de floresta semidecídua preservada (VN), de aproximadamente 5 ha, onde são observadas espécies de Cerrado e Floresta, o que caracteriza uma área transicional (Duarte et al., 2014), como referencial da condição original do solo da região. No centro da área foram retiradas quatro amostras de solo, ao longo de um transecto, equidistantes de cinco metros. Em cada ponto de amostragem foram retiradas três subamostras, que foram misturadas, formando uma amostra composta.
O solo foi preparado com duas gradagens simultâneas. O milho (cv. BR 106) foi semeado manualmente com plantadeira tipo matraca, no espaçamento de 1 metro entre linhas e cinco sementes por metro linear. As leguminosas foram semeadas 30 dias após a semeadura do milho, nas entrelinhas da cultura, sendo mantidos até o final do ciclo da cultura comercial. Aos 135 dias após a semeadura foi realizada a colheita do milho.
Após a colheita do milho foi realizada amostragem de solo nas camadas de 0,0-0,10 m e 0,10-0,20 m, nas entrelinhas da cultura principal e da espécie utilizada como adubo verde. As amostras foram acondicionadas em sacos plásticos e armazenadas em ambiente com temperatura, a 4oC, até a realização das análises. As amostras foram destorroadas, peneiradas (malha de 2 mm), homogeneizadas, retirando-se as raízes e os resíduos visíveis de plantas e animais do solo e a umidade ajustada para 40 a 60% da capacidade de campo.
Para a análise do carbono da biomassa microbiana (C-BMS) foi utilizado o método da fumigação-extração, proposto por Vance et al. (1987). Determinou-se, ainda, a respiração basal (C-CO2), obtida pela incubação das amostras com captura de CO2 em NaOH, durante sete dias, pela adaptação do método da fumigação-incubação, proposto por Jenkinson e Powlson (1976). O quociente metabólico (qCO2) foi obtido a partir da relação C-CO2/C-BMS (Anderson e Domsch, 1990) e o quociente microbiano (qMIC), pela relação C-BMS/ C-orgânico total. O conteúdo do carbono orgânico foi determinado, conforme a metodologia descrita por Claessen (1997).
Os resultados obtidos foram submetidos à análise de variância e as médias comparadas pelo teste de Tukey, a 5% de probabilidade. As análises estatísticas foram processadas por meio do programa Assistat (Silva e Azevedo, 2016).
Os parâmetros microbiológicos também foram submetidos à análise de agrupamento ("cluster analysis"), adotando-se o método do vizinho mais distante ("complete linkage"), a partir da Distância Euclidiana, para avaliar a dissimilaridade entre os sistemas de consórcio estudados.
RESULTADOS E DISCUSSÃO
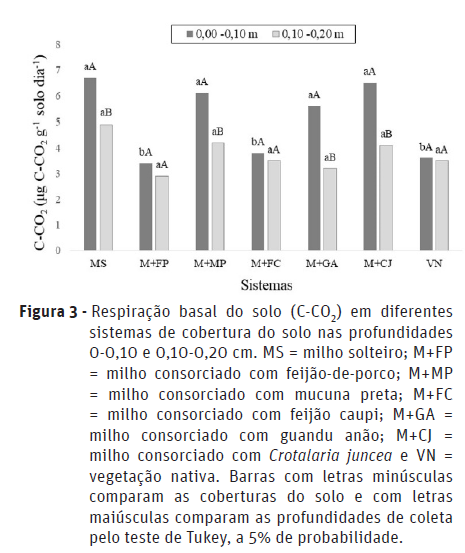
Houve interação significativa entre os fatores (consórcio x profundidade) para as variáveis carbono da biomassa microbiana (C-BMS) e respiração basal (C-CO2) (Figuras 2 e 3).
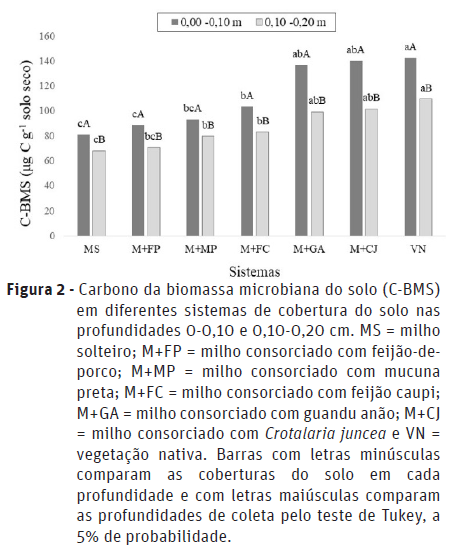
Em todos os sistemas de consórcio e no milho solteiro verificou-se que com o aumento da profundidade de amostragem houve decréscimo nos valores de C-BMS (Figura 2), indicando uma condição mais favorável a microbiota na camada mais superficial do solo. Este resultado corrobora com os obtidos por outros autores (Silva et al., 2010; Melo et al., 2017). Segundo Silva e Mendonça (2007), existe uma tendência de a biomassa microbiana do solo ser mais elevada na camada superficial do solo, devido a maior presença de matéria orgânica, água e nutrientes. Valores elevados de C-BMS indicam que os nutrientes ficam imobilizados temporariamente, o que resulta em menores perdas destes nutrientes no sistema solo-planta (Roscoe et al., 2006).
Ao comparar os sistemas de uso do solo, observou-se que a vegetação nativa (VN) foi superior em relação ao milho solteiro (MS) e aos consórcios M+MP, M+FP e M+FC, sem diferir estatisticamente do M+GA e M+CJ, independente da profundidade do solo (Figura 2). De acordo com Alves et al. (2011), os sistemas de cultivo geralmente tendem a apresentar menores teores de carbono microbiano em relação a um ambiente natural, que por sua vez, condiciona menor variação e níveis mais adequados de temperatura e umidade (Silva et al., 2010). A diversidade de espécies vegetais na mata nativa implica em deposição na serapilheira de substratos orgânicos oxidáveis com composição variada, além disso, existe maior diversidade de compostos orgânicos depositados na rizosfera, o que constitui fator favorável à biomassa microbiana (Cunha et al., 2012). Portanto, as condições distintas do solo sob vegetação de mata, juntamente com a ausência de perturbações decorrentes de atividade antrópica, tornam possível a existência de maiores quantidades de C-BMS, indicando o maior equilíbrio da microbiota do solo nesse sistema (Ferreira et al., 2010; Cunha et al., 2012; Pragana et al., 2012; Duarte et al., 2014).
No que se refere aos consórcios, o solo sob cultivo de milho com guandu-anão e com crotalária apresentou conteúdo de C-BMS muito próximo ao verificado na VN, nas duas profundidades de amostragem, com 136,6 μg C g-1 solo seco (M+GA); 139,8 μg C g-1 solo seco (M+CJ) e 142,4 μg C g-1 solo seco (VN), na camada superficial (0-0,10 m), e, 99,0 μg C g-1 solo seco (M+GA); 101, 4 μg C g-1 solo seco (M+CJ) e 109,2 μg C g-1 solo seco (VN) na camada mais profunda do solo (0,10-0,20 m) (Figura 2). Portanto, pode-se inferir que a presença dessas leguminosas no sistema provocam mudanças positivas nos teores de carbono da biomassa microbiana, assemelhando-se ao ambiente sem intervenção antrópica (condição natural). Na camada superficial, o guandu-anão e a Crotalaria juncea apresentaram incremento de C-BMS de 40,93 % e 42,27 %, respectivamente, em relação ao milho em monocultivo. Já na camada mais profunda os incrementos em relação ao MS foram de 31,62 % e 32,27 % para M+GA e M+CJ, respectivamente, demonstrando que a espécie a ser utilizada no consórcio pode determinar a qualidade da biomassa microbiana.
Para respiração basal (C-CO2), as maiores liberações foram verificadas no milho solteiro e nos consórcios M+MP, M+GA e M+CJ, na profundidade de 0-0,10 m. Já na camada mais profunda de amostragem (0,10-0,20 cm) não foi verificada diferença significativa entre as coberturas avaliadas (Figura 3).
De acordo com Cunha et al. (2011), a maior liberação de CO2 geralmente está associada à maior atividade biológica, que, por sua vez, está relacionada diretamente com a quantidade de C lábil no solo; o que explica os valores mais elevados na camada superficial do solo (Souto et al., 2009). Contudo, segundo Cunha et al. (2011), a interpretação dos resultados da atividade biológica deve ser feita com critério, pois elevados valores de respiração nem sempre indicam condições desejáveis, podendo a curto prazo significar liberação de nutrientes para as plantas e, a longo prazo, perda de C orgânico do solo para a atmosfera (Parkin et al., 1996). Portanto, elevados valores de C-CO2 podem indicar tanto situações de distúrbio quanto de alto nível de produtividade do sistema (Islam e Weil, 2000).
Não houve interação significativa entre as coberturas de solo e as profundidades de amostragem para quociente microbiano (qMIC) (Figura 4). Desta forma, os efeitos desses fatores atuaram independentemente sobre a variável. Apenas na camada 0-0,10 m foi verificada diferença significativa entre as coberturas do solo, onde os sistemas M+GA, M+CJ e VN foram superiores em comparação com o MS e M+FP, sem diferir estatisticamente do M+MP e M+FC (Figura 4).
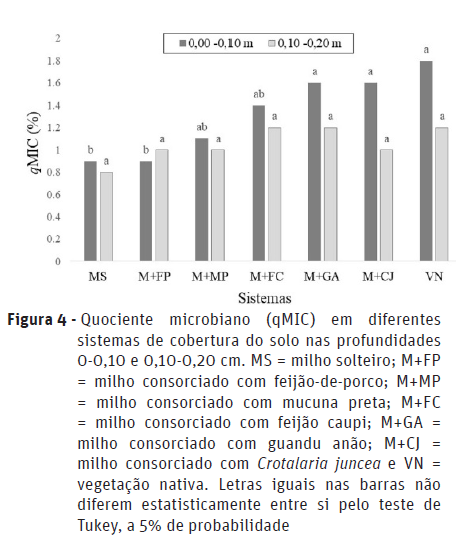
Em um estudo realizado em Santo Antônio de Goiás-GO, num Latossolo Vermelho, Cunha et al. (2011), constataram também a influência de plantas de cobertura (Crotalaria juncea; Cajanus cajan (L.) Millsp; Mucuna aterrima; Sorghum technicum e pousio) nos atributos biológicos de solo, em duas profundidades (0-0,10 m e 0,10-,0,20 m), onde verificaram qMIC variando em função das coberturas somente na camada superficial de amostragem. Este índice reflete o quanto do C orgânico está imobilizado na biomassa microbiana e mostra o potencial de reserva desse elemento no solo (Anderson e Domsch,1993), assim, valores mais elevados do quociente microbiano podem indicar um acúmulo de C no solo, enquanto valores mais reduzidos indicariam uma perda de C no solo, ao longo do tempo (Mercante et al., 2004).
Valores de quociente microbiano inferiores a 1% indicam que existe algum fator limitante à atividade microbiológica no solo (Jakelaitis et al., 2008). No presente estudo, os valores de qMIC variaram entre 0,9 % (MS e M+MP) e 1,8 % (VN) (0-0,10), estando somente os sistemas MS e M+MP com índice abaixo de 1%, evidenciando que nessas áreas pode estar ocorrendo alguma situação prejudicial a biomassa microbiana, que pode ser confirmada pelos menores teores de C-BMS (Figura 2) e maiores valores de qCO2 (Figura 5), principalmente no milho solteiro. Portanto, nos demais sistemas de consórcio e na área natural ocorreu maior incorporação do carbono orgânico do solo na biomassa microbiana, ou seja, os microrganismos imobilizaram maiores quantidades de carbono que estava disponível no solo em suas células (Dadalto et al., 2015). Na profundidade de 0,10-0,20 m houve pouca variação entre as coberturas do solo, onde apenas o MS apresentou índice inferior a 1%.
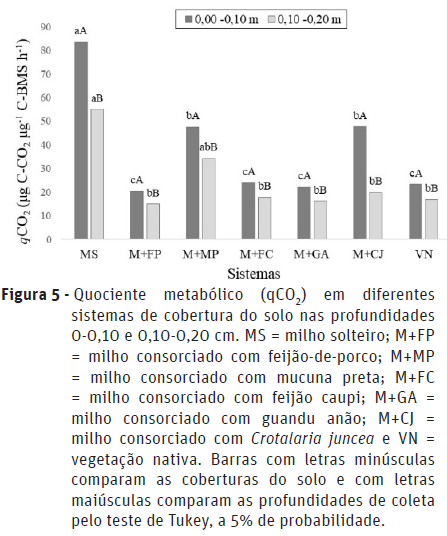
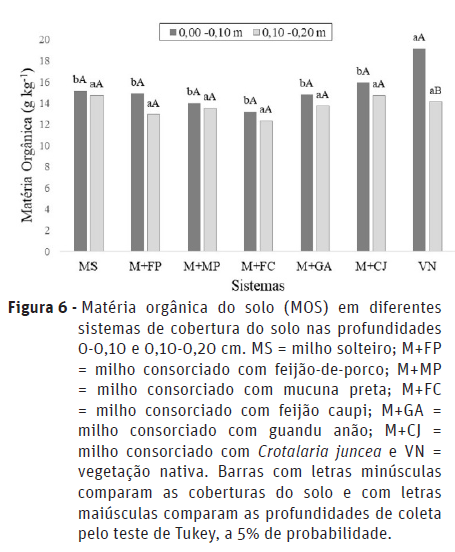
Houve interação significativa entre os fatores (consórcio x profundidade) para quociente metabólico (qCO2) e matéria orgânica do solo (MOS) (Figuras 5 e 6).
Observou-se um declínio nos valores de qCO2 em todos os sistemas de uso do solo na camada mais profunda de amostragem (0,10-0,20 m). Ao comparar os consórcios isoladamente, verificou-se que nas duas profundidades houve diferença significativa entre os sistemas (Figura 5).
Na camada mais superficial, o milho solteiro (MS) apresentou maiores valores em comparação aos sistemas de consórcio e a vegetação nativa. Já na camada mais profunda, observou-se maior qCO2 no MS em comparação ao M+FP, M+FC, M+GA, M+CJ e VN, sem diferir estatisticamente do M+MP (Figura 5).
O quociente metabólico (qCO2) é um índice que expressa a relação entre a respiração basal do solo (C-CO2) e a biomassa microbiana (C-BMS), onde valores elevados são encontrados em condições ambientais estressantes, nas quais a biomassa microbiana necessita de mais carbono para sua manutenção (Mendes et al., 2009). Portanto, reflete o influxo de energia (carbono) através da biomassa microbiana do solo (Lacerda et al., 2013). Diversos autores verificaram que a intensidade de uso e o tipo de manejo do solo em sistemas agrícolas causam aumentos no qCO2 (Silva et al., 2010; Pragana et al., 2012; Lacerda et al., 2013; Duarte et al., 2014; Gomes et al., 2015; Guimarães et al., 2017).
No presente estudo, pode-se inferir que nas duas profundidades o sistema com milho solteiro acarretou condições de estresse ou distúrbio a biomassa microbiana, em comparação aos sistemas consorciados e a vegetação nativa, que se mostraram mais equilibrados. Tal resultado pode ser decorrente de uma composição mais variada de espécies vegetais nos consórcios e VN, o que favoreceu também a maior decomposição de substratos orgânicos (Almeida et al., 2017), garantindo melhores condições para o estabelecimento da comunidade microbiana. De acordo com Tótola e Chaer (2002), um baixo qCO2 indica economia na utilização de energia e, supostamente, reflete um ambiente mais estável ou mais próximo do seu estado de equilíbrio. Ou seja, a biomassa microbiana se torna mais eficiente na utilização dos recursos do ambiente, com menor perda de carbono na forma de CO2 pela respiração e maior incorporação de carbono às células dos microrganismos (Anderson e Domsch, 1985).
Houve um declínio de MOS na vegetação nativa na camada mais profunda de amostragem do solo (Figura 6) (0,10-0,20 m), que pode estar associado com o fato de que em florestas há maior acúmulo de resíduos da parte aérea de plantas na superfície do solo, além da maior contribuição de resíduos de raízes que se reciclam em curto prazo na camada mais superficial (Karhu et al., 2011).
Ao comparar os sistemas de maneira isolada, verifica-se que somente na profundidade de 0-0,10 m houve diferença significativa entre os tratamentos (Figura 6). A VN apresentou os maiores teores de MOS em relação aos demais sistemas, que não diferiram entre si. Este resultado corrobora com os encontrados por outros autores (Duarte et al., 2014; Guimarães et al., 2017). Fontana et al. (2011), avaliando os compartimentos da matéria orgânica em solo com diferentes coberturas, verificaram menores valores de carbono orgânico total na camada mais profunda do solo, além de teores mais elevados na vegetação nativa em relação aos sistemas de cultivo; resultados próximos aos verificados no presente estudo. O maior teor de MOS na vegetação nativa deve às condições de ecossistemas naturais, onde há fornecimento constante de material orgânico mais susceptível à decomposição, permanecendo o solo coberto, com menor variação e níveis mais adequados de temperatura e umidade (Santos et al., 2004).
A ausência de diferença significativa entre os sistemas de manejo do solo (consórcios e monocultura) pode ser justificada pelo fato de que apesar da MOS ser considerada uma indicadora sensível, as alterações promovidas no solo não foram suficientes para promover modificações na sua concentração (Assis et al., 2003). Em geral, as alterações no conteúdo de MOS ocorrem em médio ou longo prazo, requerendo maior tempo para ser quantificada (Roscoe et al., 2006).
Com base na análise de agrupamento, cujo objetivo consiste em agrupar sistemas de manejo baseando-se nas características similares que estes possuem (Guimarães et al., 2017), houve a formação de dois grupos com relação aos indicadores microbiológicos. Esses grupos não apresentaram nenhuma similaridade entre si, uma vez que a sua distância de ligação foi de 100% (Figura 7).
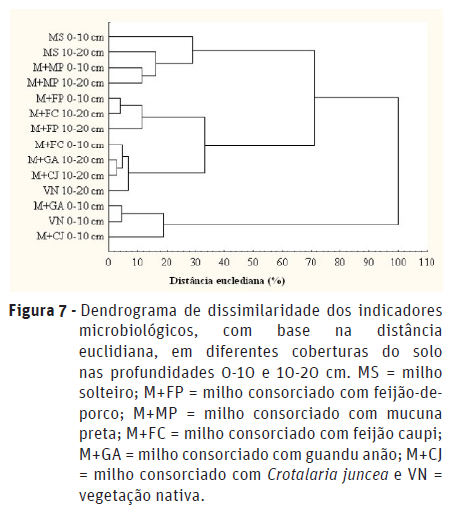
O primeiro grupo (G1) engloba os sistemas M+GA, M+CJ e vegetação nativa (VN), na menor profundidade (0-0,10 m), que apresentaram similaridade de 82 % (Figura 7). Este agrupamento ocorreu em função da semelhança entre essas coberturas do solo para as variáveis C-BMS e qMIC, evidenciando que na profundidade superficial o guandu-anão e crotalária apresenta C orgânico com qualidade semelhante ao do sistema natural.
O segundo grupo compreendeu os sistemas MS, M+MP, M+FP e M+FC, nas duas profundidades de amostragem, além do M+GA, M+CJ e VN, na camada mais profunda de avaliação (0,10-0,20 m). Neste mesmo grupo (G2) observa-se a formação de dois níveis (N1 e N2) independentes e distantes, que apresentaram dissimilaridade de 72% (Figura 7). No primeiro nível (N1) os sistemas M+FC (0-0,10 m), M+GA (0,10-0,20 m), M+CJ (0,10-0,20 m) e VN (0,10-0,20 m) se agruparam, apresentando uma semelhança de 92%. Dentro deste mesmo nível verificou-se um agrupamento constituído pelo M+FC (0-0,10 m) e M+FP, nas duas profundidades, que mostraram uma similaridade de 90%, permitindo inferir que sua distância de ligação é de 10%. Estes agrupamentos dentro do N1 apresentaram uma proximidade de 68%. Já no segundo nível (N2) verificou-se a formação de um grupo formado por MS e M+MP, nas duas profundidades de amostragem, que apresentou uma ligação de 72% de similaridade. Portanto, pode-se inferir que as modificações na biomassa microbiana do solo sob consórcio de milho e mucuna preta são próximas as do milho em cultivo solteiro.
CONCLUSÕES
A biomassa microbiana do solo e sua atividade mostraram-se sensíveis as modificações decorrentes do sistema de consórcio.
Os consórcios de milho com Crotalaria juncea e guandu-anão contribuem com o aumento do carbono da biomassa microbiana e quociente microbiano, com valores semelhantes ao do solo em condição natural, principalmente na profundidade de 0-0,10 m.
O milho em monocultivo apresenta alto quociente metabólico, pressupondo condições de estresse à comunidade microbiana do solo.
A biomassa microbiana apresenta atividade mais intensa na camada superficial do solo.
Referências bibliográficas
Almeida, A.T.; Simões, K.S.; Almeida, J.R.C.; Peixoto, C.P. & Peixoto, M.F.S.P. (2017) - Qualidade biológica do solo em sistema de policultivo no semiárido da Bahia. Ciência Agrícola, vol. 15, n. 2, p. 75-81. [ Links ]
Almeida, D.O.; Bayer, C. & Almeida, H.C. (2016) - Fauna e atributos microbiológicos de um Argissolo sob sistemas de cobertura no Sul do Brasil. Pesquisa Agropecuária Brasileira, vol. 51, n. 9, p. 1140-1147. http://dx.doi.org/10.1590/s0100-204x2016000900013 [ Links ]
Anderson, T.H. & Domsch, K.H. (1990) - Application of ecophysiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology and Biochemistry, vol. 22, n. 2, p. 251-255. https://doi.org/10.1016/0038-0717(90)90094-G [ Links ]
Anderson, T.H. & Domsch, K.H. (1985) - Determination of ecophysiological maintenance carbon requirements of soil microorganisms in a dormant state. Biology and Fertility of Soils, vol. 1, n. 2, p. 81-89. http://dx.doi.org/10.1007/BF00255134 [ Links ]
Anderson, J.P.E. & Domsch, K.H. (1993) - The metabolic quotient of CO2 (qCO2) as a specific activity paramenter to assess the effects of environmental condition, such as pH, on the microbial of forest soil. Soil Biology and Biochemistry, vol. 25, n. 3, p. 393-395. https://doi.org/10.1016/0038-0717(93)90140-7 [ Links ]
Alves, T.D.S.; Campos, L.L.; Elias Neto, N.; Matsuoka, M. & Loureiro, M.F. (2011) - Biomassa e atividade microbiana de solo sob vegetação nativa e diferentes sistemas de manejos. Acta Scientiarum. Agronomy, vol. 33, n. 2, p. 341-347. http://dx.doi.org/10.4025/actasciagron.v33i2.4841 [ Links ]
Aragão, D.V.; Carvalho, C.J.R.; Kato, O.R.; Araújo, C.M.; Santos, M.T.P. & Mourão Junior, M. (2012) - Avaliação de indicadores de qualidade do solo sob alternativas de recuperação do solo no Nordeste Paraense. Acta Amazônica, vol. 42, n. 1, p. 11-18. http://dx.doi.org/10.1590/S0044-59672012000100002 [ Links ]
Assis, É.P.M.; Cordeiro, M.A.S.; Paulino, H.B. e Carneiro, M.A.C. (2003) - Efeito da aplicação de nitrogênio na atividade microbiana e na decomposição da palhada de sorgo em solo de cerrado sob plantio direto. Pesquisa Agropecuária Tropical, vol. 33, p. 7-12. [ Links ]
Brito, M.F.; Tsujigushi, B.P.; Rocha, D.P. & Silva, R. F. (2017) - Reciclagem de nutrientes de adubos verdes e produtividade de milho cultivado em sucessão em agroecossistema de transição agroecológica. Acta Iguazu, vol. 6, n. 3, p. 11-21. [ Links ]
Claessen, M.E.C. (1997) - Manual de métodos de análise de solo. 2ª ed. Rio de Janeiro, Revista atual, Embrapa-CNPS, 212 p. [ Links ]
Cunha, E.Q.; Stone, L.F.; Ferreira, E.P.B.; Didonet, A.D.; Moreira, J.A.A. & Leandro, W. M. (2011) - Sistemas de preparo do solo e culturas de cobertura na produção orgânica de feijão e milho. II - atributos biológicos do solo. Revista Brasileira de Ciência do Solo, vol. 35, n. 2, p. 603-611. http://dx.doi.org/10.1590/S0100-06832011000200029 [ Links ]
Cunha, E.Q.; Stone, L.F.; Ferreira, E.P.B.; Didonet, A.D. & Moreira, J.A.A. (2012) - Atributos físicos, químicos e biológicos de solo sob produção orgânica impactados por sistemas de cultivo. Revista Brasileira de Engenharia Agrícola e Ambiental, vol. 16, n. 1, p. 56-63. http://dx.doi.org/10.1590/S1415-43662012000100008 [ Links ]
Dadalto, J.P.; Fernandes, H.C.; Teixeira, M.M.; Cecon, P.R. & Matos, A.T. (2015) - Sistema de preparo do solo e sua influência na atividade microbiana. Engenharia Agrícola, vol. 35, n. 3, p. 506-513. http://dx.doi.org/10.1590/1809-4430-Eng.Agric.v35n3p506-513/2015 [ Links ]
Duarte, I.B.; Gallo, A.S.; Gomes, M.S.; Guimarães, N.F.; Rocha, D.P. & Silva, R.F. (2014) - Plantas de cobertura e seus efeitos na biomassa microbiana do solo. Acta Iguazu, vol. 3, n. 2, p. 150-165. [ Links ]
Ferreira, E.P.B.; Santos, H.P.; Costa, J.R.; De-Polli, H. & Rumjanek, N.G. (2010) - Microbial soil quality indicators under different crop rotations and tillage managements. Revista Ciência Agronômica, vol. 41, n. 2, p. 177-183. http://dx.doi.org/10.1590/S1806-66902010000200002 [ Links ]
Fontana, A.; Silva, C.F.D.; Pereira, M.G.; Brito, R.J.D. & Benites, V.D.M. (2011) - Avaliação dos compartimentos da matéria orgânica em área de Mata Atlântica. Acta Scientiarum. Agronomy, vol. 33, n. 3, p. 545-550. http://dx.doi.org/10.4025/actasciagron.v33i3.5169 [ Links ]
Giongo, V.; Mendes, A.M.S.; Cunha, T.J.F. & Galvão, S.R.S. (2011) - Decomposição e liberação de nutrientes de coquetéis vegetais para utilização no semiárido brasileiro. Revista Ciência Agronômica, vol. 42, n. 3, p. 611-618. http://dx.doi.org/10.1590/S1806-66902011000300006 [ Links ]
Gomes, S.S; Gomes, M.S.; Gallo, A.S.; Mercante, F.M.; Batistote, M. & Silva, R.F. (2015) - Bioindicadores de qualidade do solo cultivado com milho em sucessão a adubos verdes sob bases agroecológicas. Revista de La Facultad de Agronomia, vol. 114, n. 3, p. 30-37. [ Links ]
Guimarães, N.F.; Gallo, A.S.; Fontanetti, A.; Meneghin, S.P.; Souza, M.D.B. Morinigo, K.P.G. & Silva, R.F. Biomassa e atividade microbiana do solo em diferentes sistemas de cultivo do cafeeiro. Revista de Ciências Agrárias, vol. 40, n. 1, p. 34-44. http://dx.doi.org/10.19084/RCA16041 [ Links ]
Islam, K.R. & Weil, R.R. (2000) - Land use effects on soil quality in a tropical forest ecosystem of Bangladesh. Agriculture Ecosystems & Environment, vol. 79, n. 1, p. 9-16. https://doi.org/10.1016/S0167-8809(99)00145-0 [ Links ]
Jackson, L.E.; Calderon, F.J.; Steenwerth, K.L.; Scow, K.M. & Rolston, D.E. (2003) - Responses of soil microbial processes and community structure to tillage events and implications for soil quality. Geoderma, vol. 114, n. 3-4, p. 305-317. https://doi.org/10.1016/S0016-7061(03)00046-6 [ Links ]
Jakelaitis, A.; Silva, A.A.; Santos, J.B. & Vivian, R. (2008) - Qualidade da camada superficial de solo sob mata, pastagens e áreas cultivadas. Pesquisa Agropecuária Tropical, vol. 38, p. 118-127. [ Links ]
Jenkinson, D.S. & Ladd, J.N. (1981) - Microbial biomass in soil: measurement and turnover. In: Paul, E.A. & Ladd, J.N. (Eds.) - Soil Biology and Biochemistry. New York, Marcel Decker, p. 415-471. [ Links ]
Jenkinson, D.S. & Powlson, D.S. (1976) - The effects of biocidal treatments on metabolism in soil-V. A method for measuring soil biomass. Soil Biology and Biochemistry, vol. 8, n. 3, p. 209-213. https://doi.org/10.1016/0038-0717(76)90005-5 [ Links ]
Karhu, K.; Wall, A.; Vanhala, P.; Liski, J.; Esala, M. & Regina, K. (2011) - Effects of afforestation and deforestation on boreal soil carbon stocks - Comparison of measured C stocks with Yasso07 model results. Geoderma, vol. 164, n. 1-2, p. 33-45. https://doi.org/10.1016/j.geoderma.2011.05.008 [ Links ]
Lacerda, K.A.P.; Cordeiro, M.A.S.; Verginassi, A.; Salgado, F.H.M. Paulino, H.B. & Carneiro, M.A.C. (2013) - Organic carbon, biomass and microbial activity in an Oxisol under different management systems. Amazonian Journal of Agricultural and Environmental Sciences, vol. 56, n. 3, p. 249-254. http://dx.doi.org/10.4322/rca.2013.036 [ Links ]
Matoso, S.C.G.; Silva, A.N.; Fiorelli-Pereira, E.C.; Coleta, Q.P. & Souza, E.F.M. (2012) - Frações de carbono e nitrogênio de um Latossolo Vermelho-Amarelo distrófico sob diferentes usos na Amazônia brasileira. Acta Amazonica, vol. 42, n. 2, p. 231-240. http://dx.doi.org/10.1590/S0044-59672012000200008 [ Links ]
Melo, V.F.; Silva, D.T.; Evald, A. & Rocha, P.R.R. (2017) - Qualidade química e biológica do solo em diferentes sistemas de uso em ambiente de savana. Revista Agro@mbiente On-line, vol. 11, n. 2, p. 101-110. [ Links ]
Mendes, I.C.; Hungria, M.; Reis-Junior, F.B.; Fernandes, M.F.; Chaer, G.M.; Mercante, F.M. & Zilli, J.E. (2009) - Bioindicadores para avaliação da qualidade dos solos tropicais: utopia ou realidade? Planaltina, Embrapa Cerrados, 31 p. [ Links ]
Mercante, F.M.; Fabricio, A.C.; Machado, L.A.Z. & Silva, W.M. (2004) - Parâmetros Microbiológicos Como Indicadores de Qualidade do Solo sob Sistemas Integrados de Produção Agropecuária. Dourados, Embrapa Agropecuária Oeste, 27 p. [ Links ]
Mercante F.M.; Silva, R.F.; Otsubo, A.A. & Melhorança, A.L. (2007) - Avaliação de plantas daninhas após cultivos de mandioca sob diferentes coberturas vegetais. Ensaios e Ciências: Ciências Biológicas, Agrárias e da Saúde, vol. 11, n. 1, p. 33-40. [ Links ]
Parkin, T.B.; Doran, J.W. & Francop-Vizcaino, E. (1996) - Field and laboratory tests of soil respiration. In: Doran, J.W. & Jones, A. (Eds) - Method for assessing soil quality. Madison, Soil Science Society of America, p. 231-245. [ Links ]
Pragana, R.B.; Nóbrega, R.S.A.; Ribeiro, M.R. & Lustosa Filho, J.F. (2012) - Atributos biológicos e dinâmica da matéria orgânica em Latossolos Amarelos na região do Cerrado piauiense sob sistema plantio direto. Revista Brasileira de Ciência do Solo, vol. 36, n. 3, p. 851-858. http://dx.doi.org/10.1590/S0100-06832012000300015 [ Links ]
Roscoe, R.; Mercante, F.M.; Mendes, I.D.C.; Reis Júnior, F.B.; Santos, J.C.F. & Hungria, M. (2006) - Biomassa microbiana do solo: fração mais ativa da matéria orgânica. In: Roscoe, R.; Mercante, F.M. & Salton, J.C. (Eds.) - Dinâmica da matéria orgânica do solo em sistemas conservacionistas: modelagem matemática e métodos auxiliares. Dourados, Embrapa, p. 163-198. [ Links ]
Santos, V.B.; Castilhos, D.D.; Castilhos, R.M.V.; Pauletto, E.A.; Gomes, A.S. & Silva, D.G. (2004) - Biomassa, atividade microbiana e teores de carbono e nitrogênio totais de um planossolo sob diferentes sistemas de manejo. Revista Brasileira de Agrociência, vol. 10, n. 3, p. 333-338. [ Links ]
Silva, E.F.; Lourente, E.R.P.; Marchetti, M.E.; Mercante, F.M.; Ferreira, A.K.T. & Fujji, G.C. (2011) - Frações lábeis e recalcitrantes da matéria orgânica em solos sob integração lavoura-pecuária. Pesquisa Agropecuária Brasileira, vol. 46, n. 10, p. 1321-1331. http://dx.doi.org/10.1590/S0100-204X2011001000028 [ Links ]
Silva, F.A.S. & Azevedo, C.A.V. (2016) - Comparison of means of agricultural experimentation data through different tests using the software Assistat. African Journal of Agricultural Research, vol. 11, n. 37, p. 3527-3531. https://doi.org/10.5897/AJAR2016.11523 [ Links ]
Silva, I.R. & Mendonça, E.S. (2007) - Matéria orgânica do solo. In: Novais, G.N.; Alvarez V., V.H.; Barros, N.F.; Fontes, R.L.F.; Cantarutti, B. & Neves, J.C.L. (Eds) - Fertilidade do solo. Viçosa, Sociedade Brasileira de Ciência do Solo, p. 275-374. [ Links ]
Silva, R.R.D.; Silva, M.L.N.; Cardoso, E.L.; Moreira, F.M.D.S.; Curi, N. & Alovisi, A.M.T. (2010) - Biomassa e atividade microbiana em solo sob diferentes sistemas de manejo na região fisiográfica Campos das Vertentes - MG. Revista Brasileira de Ciência do Solo, vol. 34, n. 5, p. 1585-1592. http://dx.doi.org/10.1590/S0100-06832010000500011 [ Links ]
Six, J.; Frey, S.D.; Thies, R.K. & Batten, K.M. (2006) - Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Science Society of America Journal, vol. 70, n. 2, p. 555-569. http://dx.doi.org/10.2136/sssaj2004.0347 [ Links ]
Song, Y.N.; Zhang, F.S.; Marschner, P.; Fan, F.L.; Gao, H.M.; Bao, X.G.; Sun, J.H. & Li, L. (2007) - Effect of intercropping on crop yield and chemical and microbiological properties in rhizosphere of wheat (Triticum aestivum L.), maize (Zea mays L.), and faba bean (Vicia faba L.). Biology and Fertility of Soils, vol. 43, n. 5, p. 565-574. http://dx.doi.org/10.1007/s00374-006-0139-9 [ Links ]
Souto, P.C.; Bakke, I.A.; Souto, J.S. & Oliveira, V.M. (2009) - Cinética da respiração edáfica em dois ambientes distintos no semi-árido da Paraíba, Brasil. Revista Caatinga, vol. 22, n. 3, p. 52-58. [ Links ]
Souza, K.B.; Pedrotti, A.; Resende, S.C.; Santos, H.M.T.; Menezes, M.M.G. & Santos, L.A.M. (2008) - Importância de Novas Espécies de Plantas de Cobertura de Solo para os Tabuleiros Costeiros. Revista da Fapese, vol. 4, n. 2, p. 131-140. [ Links ]
Tótola, M.R. & Chaer, G.M. (2002) - Microrganismos e processos microbiológicos como indicadores da qua6lidade do solo. In: Alvarez Venegas, V.H.; Schaefer, C.E.G.R.; Barros, N.F. de; Mello, J.W.V. de Costa, L.M. (Eds.) - Tópicos em ciência do solo. Viçosa, Sociedade Brasileira de Ciência do Solo, p. 195-276. [ Links ]
Vance, E.D.; Brookes, P.C. & Jenkinson, D.S. (1987) - An extraction method for measuring microbial biomass C. Soil Biology and Biochemistry, vol. 19, n. 6, p. 703-707. https://doi.org/10.1016/0038-0717(87)90052-6 [ Links ]
Wutke, E.B.; Calegari, A. & Wildner, L. P. (2014) - Espécies de adubos verdes e plantas de cobertura e recomendações para seu uso. In: Lima Filho, O.F.L.; Ambrosano, E.J.; Rossi, F. & Carlos, J.A.D.C. (Eds) - Adubação verde e plantas de cobertura no Brasil. Brasília, Empresa Brasileira de Pesquisa Agropecuária, p. 59-168. [ Links ]
Recebido/received: 2018.11.17
Aceite/accepted: 2019.02.13

