










Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Ciências Agrárias
Print version ISSN 0871-018X
Rev. de Ciências Agrárias vol.41 no.3 Lisboa Sept. 2018
https://doi.org/10.19084/RCA18084
ARTIGO
Nutrição e crescimento do tomateiro em função da interação ácido húmico-boro
Tomato nutrition and growth as related to the humic acid-boron interaction in contrasting soils
Bruno Paulo Moschini1,* e Carlos Alberto Silva2
1 Departamento de Solos/ Universidade de São Paulo, USP, Piracicaba-SP, Brasil
2 Departamento de Ciência do Solo/ Universidade Federal de Lavras, UFLA, Lavras-MG, Brasil
(*E-mail: bruno_moschini@hotmail.com)
RESUMO
O objetivo desse estudo foi avaliar o efeito de concentrações de carbono-ácido húmico (C-AH) e de suas interações com o boro (B) sobre a nutrição e o crescimento do tomateiro. O delineamento experimental utilizado foi o inteiramente casualizado, em esquema fatorial duplo 2x5, compreendendo duas doses de B, sem adição, para manter o teor atual de B, e a que visou elevar o teor de B disponível no solo a 2 mg kg-1, combinadas com cinco concentrações de C-AH (0, 5, 10, 50 e 100 mg kg-1 de solo), em três repetições. Foram avaliadas a matéria seca (MS) da parte aérea (PA), raízes (R) e total (PA+R), bem como a relação R/PA, além da disponibilidade de B no solo e no tecido vegetal. A textura e o teor de MO são fatores que modulam a produção de MS e o efeito das concentrações de C-AH sobre o tomateiro. Independentemente do solo avaliado, na ausência da adição de B, há menor crescimento e o acréscimo na concentração de C-AH reduz a produção de MS da cultura. Com a adição do B no solo, há acréscimo na MS, principalmente de raízes, à medida que aumentam as concentrações de C-AH.
Palavras-chave: acúmulo de nutrientes, adsorção, matéria orgânica, sistema radicular.
ABSTRACT
The objective of this study was to evaluate the effect of carbon-humic acid rates (C-HA) and their interaction with soil available B contents on the nutrition and growth of tomato. The experimental design adopted was a completely randomized blocks in a 2x5 factorial scheme, with two rates of B, a treatment without B to maintain the current level of available B in soil, and a treatment with the addition of B aiming to raise its level in soil to 2 mg kg-1. Boron rates in soil were combined with five concentrations of C-HA (0, 5, 10, 50 and 100 mg kg-1 of soil), with three replicates. Shoot (S) and root (R), and total (S+R) dry matter (DM) were analyzed as well as the ratio R/S, besides the availability of B in soil and plant tissue. Soil texture and OM content are factors that modulate the dry matter production and the effect of C-HA concentrations on the tomato growth. Regardless of the evaluated soil, in the treatment without B the tomato growth was lower than other treatments and the tomato DM decreased when increased the C-HA concentration. The B treatments increased DM (especially on the roots) when the C-HA concentration was increased.
Keywords: adsorption, nutrient accumulation, organic matter, root system.
INTRODUÇÃO
A disponibilidade de B para as plantas depende de sua concentração na solução do solo e dos processos associados à precipitação, lixiviação e a interação do nutriente com os colóides orgânicos e minerais do solo (Goldberg, 1997). No solo, há adsorção de B aos óxidos de ferro e de alumínio, matéria orgânica (MO) e às frações húmicas, de modo que a adsorção do nutriente aumenta com o acréscimo do pH e com a quantidade e tipo de MO presente no solo (Goldberg e Glaubig, 1985). A maior parte do B disponível no solo está ligada à fração orgânica, que retém o nutriente em ligações de alta energia (Rosolem e Bíscaro, 2007). Com o aumento do teor de MO, há maior capacidade do solo em adsorver B, demostrando que as frações orgânicas constituem-se em matrizes de adsorção/retenção do elemento químico (Azevedo et al., 2001). No entanto, o papel do AH na retenção de B nos solos parece ser altamente dependente do tipo de solo envolvido.
Em relação a interação do ácido húmico-boro, Mezuman e Keren (1981) constataram que, em solos com pH na faixa de 7,0 e com baixos teores de MO (menores que 5%), a contribuição do AH para a adsorção de B é menor, especialmente quando significativas quantidades de argila e sesquióxidos estão presentes na amostra de solo. Reforça-se a hipótese levantada com o trabalho de Gu e Lowe (1990) que aferiram que a contribuição do AH para a adsorção de B, possivelmentre, seja menor quando solos ácidos estão envolvidos. Somente em solos com altos teores de MO e pH elevado (faixa alcalina), o papel do AH pode ser dominante, com maior adsorção de B pelos óxidos e pela MO (Evans, 1987). Assim, o pH do solo que pode ser alterado pelas concentrações de C-AH (devido à natureza alcalina dos AH), pode ter um efeito positivo ou antagônico sobre a disponibilidade e dinâmica do B no sistema solo-planta. Com o aumento do pH do solo pode haver mais B adorvido nos coloides dos solos, reduzindo o B prontamente disponível às plantas.
Além de atuar sobre as formas e dinâmica do B em solo, as substâncias húmicas podem regular o crescimento e desenvolvimento das plantas (Chen e Aviad, 1990; Nardi et al., 2002). Os AHs podem interferir no metabolismo vegetal, indiretamente, por afetar as características do solo (aumento da capacidade de troca catiônica, formação de complexos organo-metálicos, aumento da absorção de nutrientes e melhoria da estrutura do solo), ou diretamente, por aumentar a produtividade e qualidade dos produtos colhidos e estimular vários processos fisiológicos e bioquímicos que promovem maior absorção de nutrientes e crescimento vegetal (Canellas et al., 2008). Os efeitos mais relatados dos AHs nas plantas estão relacionados com o maior crescimento do sistema radicular, envolvendo a formação de raízes laterais, raízes adventícias, alongamento celular e a formação de pelos radiculares, sendo esses efeitos dependentes da concentração de AH aplicada (Mora et al., 2012; Rose et al., 2014).
As ações da aplicação de AH e outras frações orgânicas sobre o crescimento e a nutrição vegetal já foi objeto de ampla revisão (Rose et al., 2014). A textura do solo, o teor de MO e a disponibilidade de B, são importantes atributos que regulam o crescimento do tomateiro. Nesse contexto, a título de hipótese, é possível antever que a aplicação de C-AH exerça efeito mais pronunciado nas raízes do que na parte aérea, por esse órgão da planta estar mais exposto ao efeito do C-AH aplicado no solo. Já as respostas para a parte aérea dependem de diversas variáveis (espécie vegetal, variedade da planta, origem e grupos funcionais presentes no AH e sua reatividade e interação com o B). No Brasil, há escassez de trabalhos que investiguem a interação ácido húmico-boro e seus efeitos na nutrição e crescimento do tomateiro, sendo a maioria dos estudos realizados em solução nutritiva e para outras culturas. Nesse sentido, objetivou-se avaliar a nutrição e crescimento do tomateiro em função das concentrações de C-AH e de suas interações com o B em solos contrastantes quanto à textura e teor de MO.
MATERIAL E MÉTODOS
O trabalho foi conduzido no ano de 2015 em casa de vegetação (estufa) no município de Lavras, Estado de Minas Gerais, Brasil (21°1334S, 44°5845W, 943 m de altitude). O clima do local é classificado como mesotérmico com estação seca de inverno e verão chuvoso (Cwb), segundo Köppen (1948). Os solos elencados para o estudo foi o Latossolo Vermelho Amarelo de textura média (LVA) e Latossolo Vermelho de textura argilosa (LV) (EMBRAPA, 2013), referentes aos Ferralsols, segundo a base de referência mundial para recursos de solos (IUSS Working Group WRB, 2015), amostrados nos municípios de Itumirim e Lavras, Estado de Minas Gerais, Brasil, respectivamente.
Antes do plantio do tomate foi realizada a amostragem de solo para caracterizar as propriedades químicas e físicas do solo sob condições naturais, a partir da coleta de três amostras simples da camada 0-20 cm (Quadro 1). Após homogeneização das amostras simples, as amostras compostas de solo foram analisadas, segundo metodologia descrita por Raij et al. (2001).
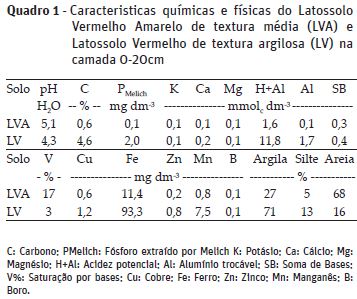
As adubações para cultivo do tomateiro em vaso foram realizadas de acordo com Novais et al. (1991), a partir da caracterização química inicial dos solos. Dessa forma, foram aplicados 4,90 e 0,63 g vaso-1 de carbonato de cálcio e 1,02 e 0,13 g vaso-1 de magnésio (proporção 3:1) no LV e LVA, respectivamente, visando elevar a saturação de bases (V%) a 65%. Após a mistura com o corretivo de acidez, os solos foram incubados por 20 dias, mantendo-se, durante esse período, a umidade próxima a 70% da capacidade de retenção máxima de água.
A adubação com P foi realizada utilizando-se duas fontes do nutriente, sendo 13% do P suprido com fosfato de potássio (K3PO4) e 87% com fosfato de amônio (NH4H2PO4), visando adicionar 200 mg kg-1 de P no LVA e 400 mg kg-1 de P no LV. A adubação com K foi realizada em ambos os solos, visando adicionar 300 mg kg-1 de K, sendo 100 mg kg-1 de K aplicadas no plantio, com a adição de 0,21 g vaso-1 de K3PO4, seguido de duas adubações de cobertura com cloreto de potássio (KCl), respectivamente, aos 10 e 20 dias após o plantio. O S foi adicionado ao solo por meio da aplicação de 0,24 g vaso-1 de sulfato de amônio [(NH4)2SO4], visando adicionar 50 mg kg-1 de S. A adubação com N visou a adição de 300 mg kg-1 de N que foi adicionada de forma distinta devido ao balanceamento necessário para cada adubação, sendo adicionado no plantio 164,92 mg kg-1 de N no LVA e 246,2 mg kg-1 de N no LV, com a adição de 0,99 g vaso-1 e 1,61 g vaso-1 de NH4H2PO4, respectivamente, e 0,24 g vaso-1 de (NH4)2SO4, seguido de duas adubações de cobertura com nitrato de amônio (NH4NO3), aos 10 e 20 dias após o plantio. As adubações com micronutrientes visaram a adição de 6 mg kg-1 de Zn, 5 mg kg-1 de Mn, 4 mg kg-1 de Fe, 2 mg kg-1 de Cu, utilizando-se as seguintes fontes: sulfato de zinco (ZnSO4.7H2O), cloreto de manganês (MnCl2.4H2O), sulfato de ferro (FeSO4.7H2O) e sulfato de cobre (CuSO4.5H2O), respectivamente.
Sementes de tomate (Solanum lycopersicum L.) da cultivar Santa Clara foram propagadas em substrato comercial em bandeja de poliestireno com duzentas células. Após o período de germinação e tendo atingido tamanho de aproximadamente 10 cm, as mudas foram transferidas para vasos plásticos, mantendo-se duas plantas por vaso, que foram preenchidos com 1,2 kg de solo cada. O delineamento experimental utilizado foi o inteiramente casualizado, em esquema fatorial duplo 2x5, compreendendo duas concentrações de B: sem adição do nutriente, para manter o teor natural de B disponível nos solos, e a adubação com o nutriente que visou elevar o teor de B a 2 mg kg-1 no solo, combinadas com quatro concentrações de AH (5, 10, 50 e 100 mg C-AH kg-1 de solo), além de um controle (solo cultivado sem adição de material húmico), em três repetições.
Como fonte de B, foi utilizado o ácido bórico (p.a.) (H3BO3), sendo adicionado em cada vaso 0,013 g de H3BO3, nos tratamentos com adição de B. O AH utilizado foi extraído de Leonardita com solução de hidróxido de potássio (KOH). O AH apresentava as seguintes características: pHágua: 9,7; CE: 10,2 mS cm-1; C: 37%; Ntotal: 6 g kg-1; P: 0,1 g kg-1; K: 42,8 g kg-1; Ca: 0,7 g kg-1; Mg: 2,3 g kg-1; S: 4,1 g kg-1; B: 84,7 mg kg-1; Cu: 5,9 mg kg-1; Fe: 2560 mg kg-1; Mn: 20,6 mg kg-1 e Zn: 75,3 mg kg-1. As concentrações de AH testadas foram definidas em relação ao teor de C presente no AH-Leonardita.
Após 30 dias de cultivo, foi amostrado o solo das unidades experimentais para realização da análise química para avaliar a disponibilidade de B. Posteriormente, as plantas foram colhidas, lavadas em água deionizada e subdivididas em parte aérea (PA) e raízes (R). Todo o material foi acondicionado em saco de papel tipo kraft e seco em estufa com ventilação forçada de ar a 65 °C até massa constante, conforme descrito por Mills e Jones Junior (1996). Em seguida, foi mensurada a produção de MS da PA, R e total (PA+R) com o auxílio de uma balança de precisão e os resultados expressos em g vaso-1. O material vegetal seco foi triturado em moinho tipo Wiley, homogeneizado, subamostrado e submetido à determinação das concentrações de macro e micronutrientes (Malavolta et al., 1997).
Os resultados foram submetidos à análise de variância (ANOVA), utilizando-se o teste F, enquanto que as médias foram comparadas pelo teste de Tukey, ambos a 5% de probabilidade e, quando significativos, foi realizada a análise de regressão polinomial de primeiro e segundo grau. Para a realização das análises estatísticas, foi utilizado o programa computacional SISVAR (Ferreira, 2011).
RESULTADOS E DISCUSSÃO
As concentrações de C-AH podem regular os teores de B disponíveis no solo em função da adição ou não de B (Figura 1). Em ambos os solos utilizados neste estudo, o aumento da concentração de C-AH diminuiu a disponibilidade de B, quando houve a adição do nutriente. Com o acréscimo da concentração de C-AH, houve redução nos teores de 0,39 e 0,76 mg dm-3 de B, representando uma redução de 22 e 39% no LV e LVA, respectivamente. Na ausência de adição de B, em ambos os solos, as concentrações de C-AH não interferiram na disponibilidade de B para as plantas. Entre as hipóteses que podem ser levantadas em relação a redução da disponibilidade do B no solo, destacam-se: (a) a extração de B do solo é feita com água quente e pode ter ocorrido a formação de um complexo ácido húmico-boro, sendo questionável a sua natureza e solubilidade em água; (b) o aumento da concentração de C-AH pode ter acrescido o pH do solo e, com isso, mais B é adsorvido aos coloides do solo, reduzindo a sua disponibilidade no meio de cultivo e (c) o B presente no solo pode ter interagido fortemente com a matriz do ácido húmico, a ponto de não ficar disponível para o tomateiro. De qualquer forma, é preciso investigar se a redução de B no solo, pela provável complexação ao ácido húmico adicionado, implica em menor absorção do nutriente pelo tomateiro, e se a magnitude desse possível efeito negativo é maior em solos mais arenosos, onde a interação negativa do ácido húmico-boro foi mais pronunciada.
A produção de MS do tomateiro bem como a relação da MS da R/PA diferiram entre as concentrações de C-AH, com a adição ou não de B no LV (Figura 2). Com o aumento das concentrações de C-AH e da disponibilidade de B no solo, houve maior produção de MS total e da PA do tomateiro, sendo que o teor natural disponível no solo (sem adição de B), em função das concentrações de C-AH, não foi capaz de promover incrementos de produção de MS. A MS de R respondeu de modo diferenciado, ou seja, reduziu com o aumento da concentração de C-AH e com a adição de B, sendo que a máxima produção de MS (2,42 g vaso-1) ocorreu com a concentração de 42,16 mg kg-1 e houve maior produção de MS a partir da concentração de 50 mg kg-1, quando não houve adição de B no solo. A relação R/PA é um atributo muito plástico e responsivo aos tratamentos que afetam a raiz, portanto, é possível observar que o tomateiro respondeu de modo diferenciado em relação à concentração de C-AH, bem como em função da disponibilidade de B no solo. Quando as condições de crescimento de raíz são ótimas, é comum que a relação R/PA seja minima, pois as plantas consomem menos fotoassimilados para absorver água e nutrientes, havendo mais reservas de produtos da fotossíntese para o crescimento da PA, como pode ser observado quando há o fornecimento adequado de B no solo. Assim, a maior proliferação de R pode não determinar maior crescimento ou produtividade da planta e, em situações onde há deficiências de nutrientes no solo, é comum que a planta direcione os fotoassimilados para as R, para assegurar as quantidades de nutrientes necessárias, a fim de que processos fisiológicos e bioquímicos das plantas não sejam comprometidos (Paiva & Oliveira, 2014).
O aumento das concentrações de C-AH com a adição de B diminuiu a produção de MS da R, sendo esses resultados reforçados pelos trabalhos desenvolvidos por Baldotto e Baldotto (2014), afirmando que, independentemente da fonte de MO utilizada, concentrações mais baixas de AH apresentam atividade estimulante na fisiologia vegetal. Por outro lado, concentrações elevadas podem resultar na diminuição do crescimento e do desenvolvimento das plantas. De acordo com Chen et al. (2004) as maiores respostas de crescimento das plantas são observadas com concentrações de 5-40 mg kg-1, com crescimento máximo a cerca de 20 mg kg-1 de AH, seguido por um aumento mais gradual de crescimento, quando são utilizadas concentrações entre 40-200 mg kg-1. Neste estudo, foi possível observar que a máxima produção de MS de R foi atingida com a utilização de 42,16 mg kg-1 de C-AH, o que está de acordo com o estudo realizado por Chen et al. (2004).
A interação do C-AH com o solo, com a adição do B, na produção de MS de R mostra a capacidade dos AH em estimular vários processos fisiológicos que promovem o crescimento vegetal, especialmente do sistema radicular. As substâncias húmicas (SH) ao regularem o crescimento das plantas atuam diretamente no meio de crescimento, como demonstrado em trabalhos desenvolvidos por Nardi et al. (2002) e Mora et al. (2012). Quando analisado as R do tomateiro com a adição de B, verificou-se que as concentrações de 10 e 50 mg kg-1 promoveram sistema radicular mais ramificado, em relação aos tratamentos que não receberam o B e ao controle. O maior desenvolvimento radicular para as concentrações de C-AH anteriormente citadas podem promover uma área de superfície radicular maior, o que pode ajudar a planta a explorar maior volume de solo, facilitando a adaptação em ambientes de baixa fertilidade e com restrições hidrícas (Canellas et al., 2002, 2008).
Com o objetivo de avaliar o desenvolvimento radicular com a aplicação de AH de lodo de esgoto e aplicação de vermicomposto em plantas de café e milho, Façanha et al. (2002) relataram um possível mecanismo de ação para o estímulo e desenvolvimento do sistema radicular, sinalizando a hipótese de que a H+-ATPases das membranas plasmáticas é um dos principais alvos moleculares envolvidos na ação dos AH sobre o crescimento das plantas. Essa enzima se caracteriza por fazer parte do principal sistema de transporte ativo de H+ da membrana plasmática e exerce forte efeito sobre a regulação do pH do apoplasto (Rayle e Cleland, 1992). A acidificação do apoplasto é uma pré-condição para o aumento da plasticidade da parede celular e, por conseguinte, para a elongação da célula vegetal, sendo esse fenômeno associado à ação de auxinas (Hager et al., 1991; Rayle e Cleland, 1992; Frias et al., 1996), que estimulam o crescimento de raízes e a absorção de nutrientes. Estes trabalhos são clássicos no que diz respeito ao entendimento do desenvolvimento radicular utilizando SH, contudo, mais informações sobre o papel das SHs em estimular as raízes e aperfeiçoar a absorção de nutrientes estão relatados na ampla revisão realizado por Rose et al. (2014).
A interação das concentrações de C-AH com a adição ou não de B diferiu em termos de efeito sobre a produção de MS no LVA (Figura 3). Com o aumento nas concentrações de C-AH sem adição de B no solo, houve menor produção de MS Total e da PA, sendo que quando o B foi adicionado no solo, há uma queda na produção de MS total até a conencetração de 50 mg kg-1, voltando a crescer com o aumento da concentração do C-AH. De modo similar ao verificado para o LV, a produção de MS de R também se comportou de modo diferenciado no LVA, dado que o aumento da concentração de C-AH sem adição de B propiciou máxima produção de MS (0,88 g vaso-1) com a concentração de 44 mg kg-1 e, quando adicionado o B, houve decréscimo de produção de MS até a concentração de 50 mg kg-1. No que se refere à relação R/PA, não houve diferenças estatísticas entre as médias dos tratamentos testados.
É possível observar que, no LV, a produção de MS foi maior, especialmente quando adicionou-se B no solo. A maior produção de MS pode estar associada à textura mais argilosa desse solo, bem como pelo seu maior teor de MO, o que aumenta a capacidade do solo em reter nutrientes. A maior presença de MO nesse solo pode implicar em maior disponibilidade de AH, de modo que o material húmico natural do solo pode ser suficiente para proporcionar maior crescimento às plantas. Assim, é provavel que a natureza química, a concentração de frações húmicas e disponibilidade de C na solução do solo, devido a maior disponibilidade de MO no LV, regulem os efeitos do C-AH adicionado, como já anteciparam Halpern et al. (2015).
Após o período de incubação do solo com os corretivos de acidez, houve aumento de pH em ambos os solos, que atingiu valores de 6,2 e 7,1 no LV e LVA, respectivamente. Essas diferenças nos valores de pH podem ter determinado os efeitos das concentrações de C-AH sobre o tomateiro. É bastante provável que o pH mais elevado no LVA tenha diminuido a disponibilidade de B, devido a maior adsorção do nutriente aos coloides do solo em ambiente alcalino. Com base nessas premissas, a adição de C-AH pode ter restringido ainda mais a oferta de B às plantas cultivadas no solo sem adição de H3BO3, possivelmente, devido as formas complexadas e menos solúveis do B presente nos solos e menos disponibilizadas às plantas. Do ponto de vista agronômico, o uso de C-AH só deve ocorrer em locais onde não há restrição na disponibilidade de B. Assim, em condições de escassez do nutriente no solo e em condições de pH com valores próximos da faixa alcalina, portanto maiores que os níveis recomendados para o cultivo do tomate, o uso de C-AH se mostra proibitivo, por restringir fortemente o crescimento do tomateiro.
Como comentado anteriormente, a disponibilidade de B no solo parece ser dependente do tipo de solo e das condições de fertilidade. Seguindo essa hipótese, Huettl (1976) verificou que a adsorção de B aumentou lentamente até pH 6,0 e com aumento sucessivo do pH houve maior adsorção de B pelo solo. Além do pH do solo influenciar a disponibilidade de B, o estudo desenvolvido por Goldberg e Glaubig (1985) mostrou que solos com maior quantidade de minerais de argila e sexquióxidos, também, adsorvem maior quantidade de B. Em relação ao papel do C-AH sobre a disponibilidade de B, Gu e Lowe (1990) verificaram a adsorção de B em AH, utilizando três tipos diferentes de solos, dos quais foram isolados e extraídos amostras distintas de AH. Os autores observaram que a adsorção de B pelo AH foi altamente dependente do pH, ocorrendo maior adsorção de B em pH 8,8 do que em pH 6,7, sendo que o aumento gradativo do pH aumentou a adsorção de B até pH 9,5.
Lemarchand et al. (2005) verificou que há menor adsorção de B pelos AH em uma faixa de pH abaixo de 6,5, o que poderia estar associado à presença da grande quantidade de cálcio disponível que pode ser utilizado para precipitar o AH ao invés do B. Segundo o autor, a maior parte dos grupos carboxílicos dos AH estariam ocupados por cálcio, por conseguinte, haveria menos sítios de adsorção no AH para a complexação do B. A suposição é que o cálcio tenha maior afinidade do que o H3BO3 para se ligar aos grupos carboxílicos dos AHs. Embora haja falta de informação para permitir uma comparação direta sobre a estabilidade do complexo ácido húmico-boro e ácido húmico-cálcio, o íon H+ presente no húmus pode reter muito mais B do que cálcio.
A concentração e acúmulo de B pelo tomateiro foi maior no LV do que nas plantas cultivadas no LVA (Figura 4). A concentração de B não diferiu entre os tratamentos, quando adicionado B no meio de cultivo, havendo apenas uma diminução discreta com o aumento das concentrações de C-AH sem adição de B no solo, no LV, sustentando a hipótese de que o uso de C-AH não deve ser recomendado quando a disponibilidade de B no solo é baixa. No LVA, o aumento da concentração de C-AH até 33,9 mg kg-1 resultou em máxima concentração de B (0,51 mg planta-1), quando o B foi adicionado ao solo. Em relação ao acúmulo de B pela PA do tomateiro, houve aumento de acúmulo no LV em função do aumento das concentrações de C-AH, e o inverso foi observado no LVA. Em ambos os solos cultivados, as concentrações de C-AH, sem a adição de B, não resultaram em diferenças no acúmulo de B pela planta. Formulou-se a hipótese que a falta de evidências mais claras pode estar associada ao método de extração, uma vez que o B é extraído com o auxílio de ácido clorídrico e azometina-H e pode acontecer de não extrair todo o B do complexo ácido humico-boro formado. Por isso, é importante testar outros métodos de extração de B, quando há maior presença de B complexado com ligantes orgânicos do ácido húmico. A combinação das concentrações de C-AH com a adição ou não de B não interferiu nas quantidades acumuladas de macro e micronutrientes no tomateiro, à exceção do B.
A aplicação de C-AH pode resultar, indiretamente, em incrementos nas concentrações e acúmulos de macro e micronutrientes pelas plantas, o que não ficou muito evidenciado neste estudo. Dessa forma, Lima et al. (2011) verificaram que a adição de AH no meio de cultivo aumentou a concentração de B em folhas de tomate, sendo que o aumento excessivo das concentrações de AH diminuiu o acúmulo do nutriente. As diferenças em relação ao acúmulo de nutrientes, referido por diversos autores, dependem da época de plantio, idade da planta, cultivar, órgão amostrado e da interação com outros nutrientes (Carvalho et al., 2004). Os resultados obtidos neste estudo sugerem que o acúmulo de B nas plantas possa estar associado com a absorção desse nutriente na forma de complexos organo-metálicos do tipo ácido húmico-boro de baixa massa molar e que podem, portanto, ser absorvidos pelas células das R e direcionados para as folhas e PA do tomateiro.
CONCLUSÕES
1. A textura do solo, matéria orgânica e a disponibilidade de B são fatores que modulam a produção de MS e o efeito das concentrações de C-AH sobre o crescimento do tomateiro.
2. O pH é outro fator que regula a ação do C-AH sobre as plantas, de modo que o pH na faixa neutra no LVA diminuiu a disponibilidade de B no solo, e isso faz com que o C-AH reduza a MS da PA do tomateiro.
3. Independentemente do solo avaliado, na ausência de B, houve menor crescimento do tomateiro e, por conseguinte, menores concentrações e acúmulo de B na PA das plantas, e o uso de concentrações crescentes de C-AH não atenua a deficiência de B no solo.
4. Em ambos os solos, com maior disponibilidade de B, foi possível obter maiores produções de MS do tomateiro à medida que se aumentavam as concentrações de C-AH.
Referências Biblográficas
Azevedo, W.R.; Faquin, V. & Fernandes, L.A. (2001) – Adsorção de boro em solos de várzea do Sul de Minas Gerais. Pesquisa Agropecuária Brasileira, vol. 36, n. 7, p. 957-964. http://dx.doi.org/10.1590/S0100-204X2001000700005 [ Links ]
Baldotto, M.A. & Baldotto, L.E.B. (2014) – Ácidos Húmicos. Revista Ceres, vol. 61, n. sp., p. 856-881. http://dx.doi.org/10.1590/0034-737x201461000011 [ Links ]
Canellas, L.P.; Mendonça, E.S.; Dobbss, L.B.; Baldotto, M.A.; Velloso, A.C.X. & Amaral Sobrinho, N.M.B. (2008) – Reações da matéria orgânica. In: Santos, G.A.; Silva, L.S. & Canellas, L.P (Eds.) – Fundamentos da matéria orgânica do solo: Ecossistemas tropicais e subtropicais. Porto Alegre-RS, Metrópole, p. 45-53. [ Links ]
Canellas, L.P.; Olivares, F.L.; Façanha, A.L.O. & Façanha, F.L. (2002) – Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiology, vol. 30, n. 4, p. 1951-1957. https://doi.org/10.1104/pp.007088 [ Links ]
Carvalho, J.G.; Bastos, A.R.R. & Alvarenga, M.A.R. (2004) – Fertirrigação. In: Alvarenga, M.A.R. (Ed.) – Tomate: produção em campo, em casa-de-vegetação e em hidroponia. Lavras-MG, Editora UFLA, p. 63-120. [ Links ]
Chen, Y.; De Nobili, M. & Aviad, T. (2004) – Stimulatory effects of humic substances on plant growth. In: Magdoff, F. & Weil, R.R. (Eds.) – Soil Organic Matter in Sustainable Agriculture. Boca Raton-FL, CRC Press, p. 103-130. [ Links ]
Chen, Y. & Aviad, T. (1990) – Effects of humic substances on plant growth. In: MacCarthy, Y.P. (Ed.) – Humic substances in soil and crop sciences: Selected readings. Chicago-IL, CAB, p.161-187. [ Links ]
EMBRAPA (2013) – Sistema brasileiro de classificação de solos. 3ª ed. Rio de Janeiro-RJ, Centro Nacional de Pesquisas de Solos, Empresa Brasileira de Pesquisa Agropecuária. 353 p. [ Links ]
Evans, L.J. (1987) – Retention of boron by agricultural soil from Ontario. Canadian Journal of Soil Science, vol. 67, n. 1, p. 33-42. https://doi.org/10.4141/cjss87-003 [ Links ]
Façanha, A.A.; Façanha, A.L.O.; Olivares, F.L.; Guridi, F.; Santos, G.A.; Velloso, A.C.X.; Rumjanek, V.M.; Brasil, F.; Schripsema, J.; Braz-Filho, R.; Oliveira, M.A. & Canellas, L.P. (2002) – Bioatividade de ácidos húmicos: efeitos sobre o desenvolvimento radicular e sobre a bomba de prótons da membrana plasmática. Pesquisa Agropecuária Brasileira, vol. 37, n. 9, p. 1301-1310. http://dx.doi.org/10.1590/S0100-204X2002000900014 [ Links ]
Ferreira, D.F. (2011) – SISVAR: um programa para análises e ensino de estatística. Ciência & Agrotecnologia, vol. 35, n. 6, p. 1036-1042. http://dx.doi.org/10.1590/S1413-70542011000600001 [ Links ]
Frias, I.; Caldeira, M.T.; Perez, J.R.C.; Avino, J.P.A.; Macia, F.A.C.; Kuppinger, O.; Stransky, H.; Pages, M.; Hager, A. & Serrano, R. (1996) – A major isoform of the maize plasma membrane H+-ATPase, characterization and induction by auxin in coleoptiles. The Plant Cell, vol. 8, n. 9, p. 1533-1544. https://doi.org/10.1105/tpc.8.9.1533 [ Links ]
Goldberg, S. & Glaubig, R.A. (1985) – Boron adsorption on aluminum and iron oxide minerals. Soil Science Society of America Journal, vol. 49, n. 6, p. 1374-1379. https://doi.org/10.2136/sssaj1985.03615995004900060009x [ Links ]
Goldberg, S. (1997) – Chemistry and mineralogy of boron in soils. In: Gupta, U.C. (Ed.) – Boron and its role in crop production. Boca Raton-FL, CRC Press, p. 3-44. [ Links ]
Gu, B. & Lowe, L.E. (1990) – Studies on Adsorption of boron on humic acids. Canadian Journal of Soil Science, vol. 70, n. 3, p. 305-311. https://doi.org/10.4141/cjss90-031 [ Links ]
Hager, A.; Debus, G.; Edel, H.G.; Stransky, H. & Serrano, R. (1991) – Auxin induces exocytosis and rapid synthesis of a high-turnover pool of plasma-membrane H+-ATPase. Planta, vol. 185, n. 4, p. 527-537. https://doi.org/10.1007/BF00202963 [ Links ]
Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T. & Yermiyahu, U. (2015) – The Use of Biostimulants for Enhancing Nutrient Uptake. Advances in Agronomy, vol. 130, p. 141–174. https://doi.org/10.1016/bs.agron.2014.10.001 [ Links ]
Huettl, P.J.V. (1976) – The pH-dependent sorption of boron by soil organic matter. M. Sc. Thesis. Madison-WI, University of Wisconsin, 127 p. [ Links ]
IUSS Working Group WRB – (2015) – World reference base for soil resources 2014. 1ª ed. Rome, Food and Agriculture Organization of the United Nations (FAO), 203 p. [ Links ]
Köppen, W. (1948) – Climatology: a study of the climates of the earth. Mexico City, Fondo de Cultura Economica, 233 p. [ Links ]
Lemarchand, E.; Schott, J. & Gaillardt, J. (2005) – Boron isotopic fractionation related to boron sorption on humic acid and the structure of surface complexes formed. Geochimica et Cosmochimica Acta, vol. 69, n. 14, p. 3519-3533. https://doi.org/10.1016/j.gca.2005.02.024 [ Links ]
Lima, A.A.; Alvarenga, M.A.R.; Rodrigues, L. & Carvalho, J.G. (2011) – Leaf nutrient content and yield of tomato grown in different substrates and doses of humic acids. Horticultura Brasileira, vol. 29, n. 1, p. 63-69. http://dx.doi.org/10.1590/S0102-05362011000100011 [ Links ]
Malavolta, E.; Vitti, G.C. & Oliveira, S.A. (1997) – Avaliação do estado nutricional das plantas: princípios e aplicações. Piracicaba-SP, POTAFOS, 319 p. [ Links ]
Mezuman, U. & Keren, R. (1981) – Boron adsorption by soils using a phenomenological adsorption equation. Soil Science Society of America Journal, vol. 45, n. 4, p. 722-726. https://doi.org/10.2136/sssaj1981.03615995004500040009x [ Links ]
Mills, H.A. & Jones Junior, B. (1996) – Plant analysis handbook II: A practical sampling, preparation, analysis and interpretation guide. 2ª ed. Athens, Micro Macro Publishing, 422 p. [ Links ]
Mora, V.; Baigorri, R.; Bacaicoa, E.; Zamarreño, A.M. & García-Mina, J.M. (2012) – The humic acid-induced changes in the root concentration of nitric oxide, IAA and ethylene do not explain the changes in root architecture caused by humic acid in cucumber. Environmental and Experimental Botany, vol. 76, p.24-32. https://doi.org/10.1016/j.envexpbot.2011.10.001 [ Links ]
Nardi, S.; Pizzeguello, D.; Muscolo, A. & Vianello, A. (2002) – Physiological effects of humic substances oh higher plants. Soil Biology & Biochemistry, vol. 34, n. 11, p. 1527-1536. https://doi.org/10.1016/S0038-0717(02)00174-8 [ Links ]
Novais, R.F.; Neves, J.C.L. & Barros, N.F. (1991) – Ensaio em ambiente controlado. In: Oliveira, A.J.; Garrido, W.E.; Araújo, J.D. & Lourenço, S. (Eds.) – Método de pesquisa em fertilidade de solos. Brasília-DF, EMBRAPA-SEA, p. 189-253. [ Links ]
Paiva, R. & Oliveira, L.M. (2014) – Bases do crescimento e desenvolvimento vegetal. In: Paiva, R. & Oliveira, L.M. (Eds.) – Fisiologia e Produção Vegetal. Lavras-MG, Editora UFLA. [ Links ]
Raij, B. van; Andrade, J.C.; Cantarella, H. & Quaggio, J.A. (2001) – Análise química para avaliação da fertilidade do solos tropicais. Campinas-SP, Instituto Agronômico, 285 p. [ Links ]
Rayle, D.L. & Cleland, R.E. (1992) – The acid growth theory of auxin-induced cell elongation is alive and well. Plant Physiology, vol. 99, n. 4, p. 1271-1274. [ Links ]
Rose, T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R. & Cavagnaro, T.R. (2014) – A meta-analysis and review of plant growth response to humic substances: practical implications for agriculture. Advances in Agronomy, vol. 124, p. 37-89. https://doi.org/10.1016/B978-0-12-800138-7.00002-4 [ Links ]
Rosolem, C.A. & Bíscaro, T. (2007) – Adsorção e lixiviação de boro em Latossolo Vermelho-Amarelo. Pesquisa Agropecuária Brasileira, vol. 42, n. 10, p. 1473-1478. http://dx.doi.org/10.1590/S0100-204X2007001000015 [ Links ]
Agradecimentos
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, processos 303899/2015-8 e 461935/2014-7), à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, PROEX-AXUPE 590/2014), por custearem as ações de pesquisa deste estudo e financiarem bolsas de pesquisa aos autores e a Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG).
Recebido/received: 2018.03.27
Recebido em versão revista/received in revised form: 2018.05.03
Aceite/accepted: 2018.05.07

