










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Ciências Agrárias
Print version ISSN 0871-018X
Rev. de Ciências Agrárias vol.41 no.2 Lisboa June 2018
https://doi.org/10.19084/RCA17297
ARTIGO
Agronomic characteristics, nutritional status and yield of corn intercropped with dwarf pigeon pea in different spatial arrangements of plants
Características agronómicas, estado nutricional e produtividade do milho em consociação com guandu-anão, em diferentes arranjos espaciais de plantas
Anderson de S. Gallo*1, Anastacia Fontanetti2, Nathalia de F. Guimarães1, Kátia P. G. Morinigo2, Maicon D. B. de Souza2 e Rogério F. da Silva3
1Department of Soils, Agronomy Institute, Federal Rural University of Rio de Janeiro, Seropédica, Rio de Janeiro, Brazil
2 Department of Rural Development, Federal University of São Carlos, Araras, São Paulo, Brazil
3State University of Mato Grosso do Sul, Glória de Dourados, Mato Grosso do Sul, Brazil
(*E-mail: andersondsgallo@hotmail.com)
ABSTRACT
Corn (Zea mays L.) is among the most produced crops in intercropping systems, mainly due to its architecture and ecophysiology. In intercropping systems, crop management should be planned to avoid interspecific competition for production factors such as water, light and nutrients. The aim of this study was to evaluate the agronomic characteristics, nutritional status and grain yield of corn intercropped with dwarf pigeon pea (Cajanus cajan L.) in different arrangements of plants. The treatments consisted of different arrangements of dwarf pigeon pea intercropped with corn. In general, the arrangements did not negatively influence the agronomic characteristics and the growth of Poaceae legume. Sowing dwarf pigeon pea in the same rows as corn and in two rows between them increased nitrogen content in corn plants, as well as provided a higher grain yield than other arrangements including the corn monoculture. The evaluated intercropping system is a promising alternative system for a more sustainable agricultural production, with less dependence on synthetic fertilizers.
Keywords: Zea mays L., Cajanus cajan L., Interspecific competition, macronutrients.
RESUMO
O milho (Zea mays L.) está entre as culturas mais produzidas em sistemas consociados, devido principalmente a sua arquitetura e ecofisiologia. Em consociação, a gestão das culturas deve ser planeada de modo a evitar competição interespecífica por fatores de produção, tais como a água, luz e nutrientes. No presente trabalho avaliaram-se algumas características agronómicas, o estado nutricional e a produtividade do milho em modo de produção em consociação com guandu-anão (Cajanus cajan L.) em diferentes arranjos de plantas. Os tratamentos consistiram de diferentes arranjos de guandu-anão em consociação com a cultura do milho. Em geral, os arranjos não influenciaram negativamente as características agronómicas e o crescimento da planta Poaceae. O tratamento com guandu-anão semeado na linha e em duas linhas na entrelinha do milho aumentou o teor de foliar azoto nas plantas de milho, e proporcionou uma maior produtividade relativamente aos outros tratamentos, incluindo o milho em monocultura. A consociação revelou-se um sistema de produção promissor numa agricultura mais sustentável, com menor dependência de fertilizantes minerais.
Palavras-chave: Zea mays L., Cajanus cajan, competição interespecífica, macronutrientes.
INTRODUCTION
Intercropping consists of cultivating two or more species in the same area for a certain period of time, even if the crops are not necessarily sown or harvested simultaneously (Zhang et al., 2015). It is recommended to be used in many parts of the world for food or fiber production, due to its high global productivity, effective pest and disease control, ecological services and economic profitability (Wu and Wu, 2014).
Despite the advantages, in these systems, interspecific interference may impair the establishment and productivity of the species (Li et al., 2011), especially when the agricultural area is inadequately managed (Zhang et al., 2007). Therefore, the knowledge of the behavior of the species regarding competition and water, light and nutrient needs is of great importance for the success of the system (Pariz et al., 2011). The competition depends on the characteristics of the species involved, their root systems and the availability of resources in the system (Casper et al., 1998; Mushagalusa et al., 2008). Moreover, allelopathy, which consists of the plant's ability to release substances to inhibit the growth or development of another nearby plant, should also be considered in the choice of species to be grown in an intercropping system (Cheng and Xu, 2013).
Corn is among the most cultivated crops in intercropping systems, especially the traditional intercropping of corn and bean, practiced by family farmers throughout Brazil. The favorable characteristics of this cereal, such as higher dry matter accumulation rate in the early stages of development and high plant height and ear insertion height, allow harvesting to occur without interference from intercropped bean (Alvarenga et al., 2006).
Recently, the interest on the intercropping of grain-producing plants with green manures has increased significantly. This system aims to produce grains and/or straw for soil cover. The production of straw is fundamental, especially in regions with high temperature and high rainfall, as these factors increase the decomposition rate, making soil cover not feasible (Gitti et al., 2012).
Green manure promotes soil protection, and chemical, physical and biological characteristics of the soil, which make it an excellent management option; especially in organic farming systems, since mineral fertilizers have high prices and are not always allowed (Benicasa et al., 2010).
The intercropping of corn with cover crops, in particular of the Fabaceae family, is among the most indicated ones (Peng et al., 2009) due to their ability to fix atmospheric nitrogen (N2) by means of symbiosis with bacteria of the genus Rhizobium, providing the largest amount of this nutrient for crops of economic interest (Wu and Wu, 2014; Zhang et al., 2015). Furthermore, corn and Fabaceae plants present complementarity in the use of production factors (Mushagalusa et al., 2008).
The dwarf pigeon pea is another plant species cultivated in intercropping with corn, because the lower production of dry matter of this Fabaceae avoids competition with the cereal for water and nutrients and does not impair harvest (Cortez et al., 2009). In addition, it is a shrub, erect, annual or semi-perennial species that exerts multiple functions in the production systems, such as nitrogen supply (Yusuf et al., 2009), nutrient recycling and disruption of compacted soil layers due to its pivotal and deep root system (Godoy et al., 2009).
Intercropping is an important practice for the sustainability of agricultural systems. Notwithstanding, the management of intercropping systems is complex and needs to be planned in order to minimize interspecific competition, which can lead to significant productivity losses, influenced by factors such as climatic conditions, soil fertility, density and spatial arrangement of plants in the system (Lithourgidis et al., 2011). Therefore, it is fundamental to carry out studies that evaluate the behavior of the intercropped species, as well as the nutritional dynamics in intercropping systems and the grain yield, in order to find out techniques able to improve this cropping (Oliveira et al., 2011).
In view of the above, the present study aimed to evaluate the agronomic characteristics, nutritional status and grain yield of corn intercropped with dwarf pigeon pea (Cajanus cajan L.) in different arrangements of plants.
MATERIALS AND METHODS
The study was conducted from December 2014 to July 2015, in an experimental area in the municipality of Araras - SP, Brazil, at the geographical coordinates 22°1756.9 S and 47°2253.80 W, and at an altitude of 701 m. The locations soil is classified as Dystrophic Red Latosol (Oxisol), of a clayey texture, with the following chemical features in the 0.0 – 0.20 m layer: pH (in CaCl2) = 5.5; extractable P = 16.5 mg kg-1; exchangeable K = 4.1 mmolc kg-1, Ca = 28.5 mmolc kg-1, Mg = 10.0 mmolc kg-1; exchangeable acidity (H + Al) = 22.0 mmolc kg-1; soil organic matter = 23.5 g kg-1, and base saturation (V) = 65.5%. The climate of the region is of the Cwa mesothermic type, according to the Köppen classification, characterized by hot and humid summers and dry winters. In Table 1, the climatic conditions observed during the course of the experiment are summarized.
The soil was prepared with a disk plough followed by a leveler. The experimental design adopted was of randomized blocks with four repetitions. The treatments consisted of dwarf pigeon pea (Cajanus cajan L.) in different arrangements, intercropped with corn (Zea mays L.) (Al-Avaré cultivar): CM – corn monoculture; PR – dwarf pigeon pea in the same row as the corn (10 pigeon pea plants m-2); P1B – one row of dwarf pigeon pea sown between the rows of corn (10 pigeon pea plants m-2); P2B – two rows of dwarf pigeon peas sown between the rows of corn (20 pigeon pea plants m-2); PR1B – dwarf pigeon pea sown in the same rows as the corn and in a row between them (20 pigeon pea plants m-2); and PR2B - dwarf pigeon pea sown in the same rows as the corn and in two rows between them (30 pigeon pea plants m-2).
The experimental plot was formed of five rows of corn, with 0.90 m spaces between them, and six seeds were sown per meter, aiming for a population of 50,000 corn plants per hectare, after thinning. The dwarf pigeon pea was sown in a density of 10 seeds per meter, in the same rows as the corn and between them, according to the treatments above. The corn and dwarf pigeon pea were sown on 17/12/2014. The three central rows of corn in each plot were considered for the evaluation.
Weed control was carried out manually in two periods, the first 20 days after the emergence of corn (DAE) (V4-V5 stage) and the second 48 days after the emergence of corn (V9-V10 stage). For fertilizing, 800 kg of dry organic compost was used, equivalent to 13 t ha-1, and distributed homogeneously over the soil, in the planting row. The organic compost used in the study presented: pH (in H2O) = 8.0; organic C = 131.0 g kg-1; organic N = 13.0 g kg-1; P = 13, 65 mg kg-1; K = 13.44 mg kg-1; Ca = 0.19 cmolc kg-1; Mg = 0.35 cmolc kg-1; S = 0.49 cmolc kg-1; Cu = 69.2 mg kg-1; Fe = 561.4 mg kg-1; Mn = 511.2 mg kg-1; Zn = 766 mg kg-1; organic matter = 22.58 % and humidity = 37.40 %.
During the experimental period, the following variables were evaluated: dry matter of shoots of corn and dwarf pigeon pea (kg ha-1); percentage of soil cover by dwarf pigeon pea and leaf area index (LAI) of corn. These were carried out in three periods (before weed control), the first being at 20 days after corn emergence (DAE) (V4-V5 stage); the second at 48 days after corn emergence (V9-V10 stage) and the third at 85 days after emergence (R3-R4 stage), except for the variable percentage of soil cover by dwarf pigeon pea, which was evaluated in two periods, the first being at 20 days after corn emergence (DAE) (V4-V5 stage) and the second at 48 days after corn emergence (V9-V10 stage). At 85 DAE, the corn stem diameter (expressed in mm) was evaluated using a digital caliper and the height of corn plants was determined (m) using a graded ruler, considering the height between the ground level and the last fully expanded leaf.
At corn harvest, the following variables were measured: first ear insertion height (FEIH); final plant stand (FPS); number of ears per plant (NEP); number of rows of grains per ear (NRE); number of grains per row in the ear (NGR), number of grains per ear (NGE); thousand grain weight (TGW) and grain yield (GY).
To determine the dry matter of shoots of corn plants and the LAI, three plants per plot were randomly collected. For the dwarf pigeon pea, a template measuring 0.25 x 0.25 m was used. The material collected was dried in an oven with forced air circulation at 65 ºC until reaching a constant mass, being subsequenlty weighed.
The percentage of soil cover by dwarf pigeon pea plants was determined by the method of number of intersections, according to the methodology described by Fávero et al. (2001). The soil cover percentage data were transformed in arcsin √x/100 to fit normal distribution.
The leaf area index (LAI cm2 leaf cm-2 soil) was obtained by collecting three corn plants in the border of each plot. The plants were immediately taken to the laboratory for evaluation. A leaf area integrator LI-COR, model LI-3000C was used. LAI was estimated by the ratio between the total leaf area and the soil space occupied by the plants. The first ear insertion height, expressed in centimeters (cm), was determined with the help of a graduated ruler in ten representative plants of the plot. The final plant stand (plants ha-1) was obtained by counting the number of total corn plants in the useful area of each plot, with the exception of broken and bedded plants.
The corn harvest was done manually, removing all the ears contained in the useful area of each plot. The moisture content of the grains was standardized to 13%, the yield being obtained as a function of the weight of grains harvested in each plot (kg ha-1). Thousand grain weight was determined by estimating the average weight of three samples of 1000 grains per plot. The number of rows of grains and number of grains per row in the ear were obtained by counting, in ten spikes chosen at random. The number of ears per plant was obtained by the ratio between the number of ears harvested and the number of plants in the useful area of the plots.
In addition, the levels of N, phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg) and sulfur (S) in the corn leaves were determined by collecting the leaf located opposite and below the upper ear in female flowering in all treatments. Ten leaves per plot were collected at 85 DAE and, after exclusion of the central vein, plant material was dried in a oven with forced air ventilation, at 65 °C for 48 h. Then, plant samples were ground in a Wiley mill and submitted to content analysis, according to the methodology described by Malavolta et al. (1989).
To interpret the corn leaf macronutrient contents, the deviation from optimal percentage (DOP) method was used, as proposed by Montañés et al. (1993). This method is defined as the standard deviation in the concentration of an element in relation to the optimal level taken as a reference value. It is obtained using the following formula:
DOP= [(C x 100/Cref) - 100]
In which C is the concentration of the nutrient in the dry matter from the sample and Cref is the optimal concentration of the nutrient in dry matter.
The values that are considered optimal for corn cultivation were proposed by Cantarella et al. (1996). As the adequate macronutrient levels for the crop are mentioned in adequate value ranges, the lowest value in the range for each macronutrient was considered.
The DOP indices were interpreted as follows: the absolute values (without sign) indicate the importance or severity of the deficiency or excess of the nutrient. The negative values indicate a situation of macronutrient deficiency; the positive values reflect situations of excess: and the indices equal to zero indicate optimal macronutrient values (Damián-Nava et al., 2006).
The results were submitted to analysis of variance (ANOVA) and the means were compared by the Tukey test, at 5% probability.
RESULTS AND DISCUSSION
For dry matter of shoots (DMS) of dwarf pigeon pea, a significant difference was observed between treatments in the first two periods of evaluation. In the third evaluation, there was no significant difference between treatments (Table 2). In the first evaluation (20 DAE), treatments PR2B, P2B and PR1B were superior in relation to PR and P1B. In these evaluation (48 DAE), P2B and PR2B were superior to the other treatments, which did not differ from each other (Table 2).
The dwarf pigeon pea showed low DMS production, a result associated with its slow initial growth, characteristic of the species. Furthermore, the low production of the dwarf pigeon pea can be attributed to the intrinsic characteristics of the plant physiology, since it presents C3-type metabolism, and is less efficient in the fixation of atmospheric carbon (C) in relation to corn, which has a C4 photosynthetic metabolism, due to the higher photorespiration and for spending more energy in the fixation of C in environments of higher temperatures due to O2 fixation (Santos et al., 2011).
Concerning the percentage of soil cover by dwarf pigeon pea plants, there was no significant difference in the first sampling period (Table 2). In the evaluation performed in the second period, there was a difference between treatments. PR2B presented higher averages compared to PR, not differing statistically from the P2B system (Table 2). This result is attributed to a larger population of dwarf pigeon pea plants in these two systems. In addition, these two treatments accounted for the highest dry matter of shoots, with 948.4 kg ha-1 (PR2B) and 886.1 kg ha-1 (P2B). Boer et al. (2008), when evaluating the green and dry matter production and the percentage of soil cover of three cover crop species, also verified a relationship between the amount of dry matter accumulated and the percentage of soil cover.
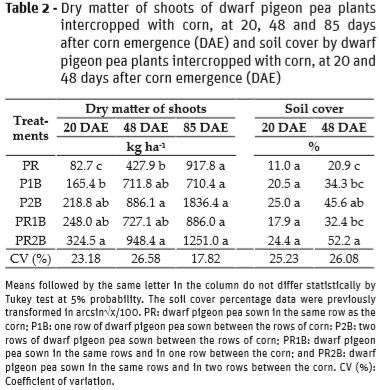
There was no significant differences between treatments with regard to the DMS of the corn plants in the three sampling periods (Figure 1). It can be inferred that the dwarf pigeon pea did not compete for production factors with corn plants, as well as there was no allelopathic effect of the dwarf pigeon pea on the Poaceae. Corn is considered a good competitor in relation to smaller plants, mainly due to its higher rate of dry matter accumulation in the early stages of development. It shows high capacity of interception of the photosynthetically active radiation along its canopy, which reduces the amount of this resource for the other species (Liu and Song, 2012).
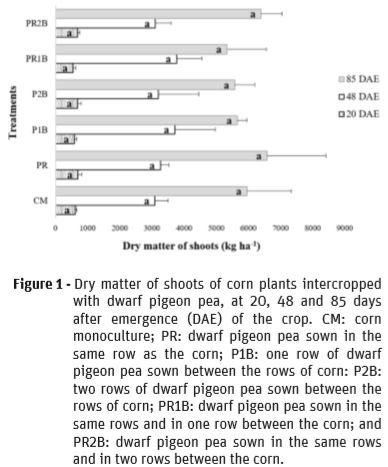
For the leaf area index (LAI) of corn, there was a significant difference between the treatments studied in the first evaluation period (Figure 2). The treatments CM, PR, P1B, P2B and PR2B were statistically similar, and superior to PR1B. This result is related to the stage of corn development, which in the initial phase shows growth from seed reserves. This growth may vary according to the size of the seed (Sangoi et al., 2004). However, Ritchie et al. (2003) mention that in the second week after emergence, photosynthesis already exerts its function in the plant and the seed reserves are depleting. Therefore, corn may have been affected by competition with dwarf pigeon pea plants for resources such as water, light and nutrients. Although corn is a great competitor, intercropping can reduce maximum LAI values (Oliveira et al., 2011).
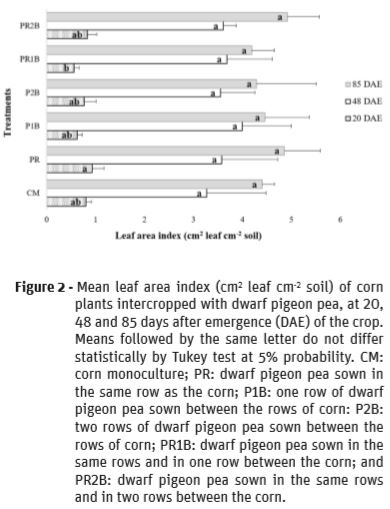
For the second and third evaluation periods, there was no significant difference between treatments for the LAI (Figure 2). The LAI data can be considered satisfactory, since values were higher than 3 in the two evaluation periods Lauer et al. (2004) referred that LAI values for corn must be between 4 and 5, at flowering, in order to optimize its agronomic performance.
For P, Mg, and S contents, no significant differences were found among treatments (Table 3). Regarding corn leaf N content, there was a significant difference between the treatments (Table 3). The PR2B arrangement was higher than the others, which did not differ between each other. The higher number of dwarf pigeon pea plants in the PR2B treatment probably caused a higher amount of N which could have been taken up by the corn plants. An increase in N availability in soil is accompanied by a positive response in N content in the leaves (Nascimento et al., 2012).
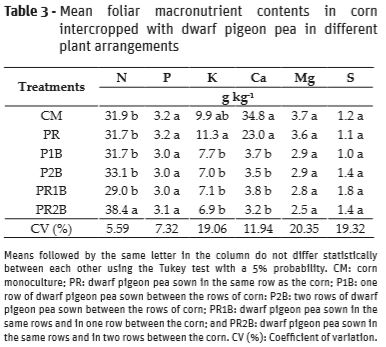
The N values obtained in all of the treatments were considered sufficient for corn, since they varied between 29.0 g kg-1 and 38.4 g kg-1. Then, monocrop or intercropping corn with dwarf pigeon pea in different plant arrangements produced foliar N content within the adequate (or slightly higher) range for corn (27.5 to 32.5 gkg-1), according to Malavolta et al. (1989). Corn behaves well in intercropping systems, and because it is an extremely demanding crop in terms of nutrients, especially N, the Fabaceae plant favored cereal growth in intercropping system.
For corn foliar K content, a significantly higher value was found under PR treatment compared to P1B, P2B, PR1B and PR2B, and did not differ statistically from MC (Table 3). The presence of dwarf pigeon pea in the different arrangements hampered the absorption of K by the corn plants, except when the Fabaceae plant was sown in the same row as the cereal. Probably, fabaceae can absorb K at depths greater than the area exploited by corn roots, and thus in the same row the species did not compete for the P available in the soil. Moreover, dwarf pigeon pea dry matter production in the rows of corn was lower than in the other arrangements, which may have reduced competition for the nutrient.
A significantly higherfoliar Ca contents were observed for CM and PR treatments (Table 3). The reduction in nutrient content in plant tissue in intercropping may have been linked to the interspecies competitive interaction exercised by one species over the other (Viera et al., 2013). The low ability to compete for Ca on the part of corn in intercropping systems was already observed by other authors (Cury et al., 2012). Higher populations of the intercropped plant make greater soil exploitation possible, intensifying the competitive effects over the main crop (Belel et al., 2014).
In relation to the rates of deviation from the optimal percentage (DOP), excessive N and P in corn leaves was verified in all of the treatments. The treatment PR2B presented the highest DOP index for N, and the CM and PR presented the highest DOP indices for P (Table 4).
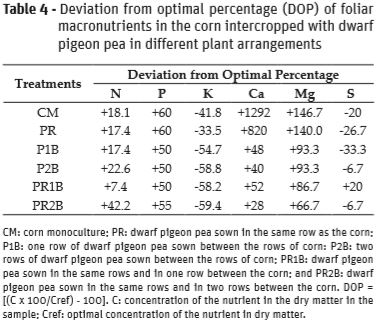
For K, all of the treatments presented negative indices, which indicates a deficiency in the macronutrient in the corn (Table 4). The K deficiency in all of the treatments may be related with the excessive Ca verified in the corn leaves (Table 4). Calcium had positive indices in all treatments, showing good availability of the nutrient in the soil, especially in the CM and PR treatments, which presented very high indices. The higher availability of Ca in present soil may cause its diffusion to roots in greater quantity, and as Ca and K are absorbed by the same mechanisms in the cellular membrane, its absorption was probably preferential in relation to K (Medeiros et al., 2008).
For Mg, positive indices were observed in all of the treatments (Table 4), showing an excess of the nutrient in the corn leaves. The highest foliar Mg indices were obtained by the CM and PR control treatments. Similar results were verified for foliar P. This similarity may be related to the synergism between these two macronutrients. According to Malavolta et al. (1989), absorption of P is influenced by the Mg concentration in the medium, with Mg able to carry P into the plant.
In relation to foliar S in the corn plants, a deficiency was verified in the CM, PR, P1B, P2B, and PR2B treatments. In the PR1B, there was an excess, even though it was low (Table 4). It is worth noting that the S deficiency occurred to a greater degree in the treatments with lower dwarf pigeon pea plant numbers and in the corn monocrop. However, greater competition for S was expected in plots with higher dwarf pigeon pea populations, since sulfur is required in the fixing nodules for symbiotic fixation of N2, given that this nutrient is an element which constitutes nitrogenase (Paiva and Nicodemo, 1994).
In the PR1B treatment, which was the only one that presented excessive S in the corn leaves, the lowest foliar content and lowest DOP index for N was verified (Tables 3 and 4). From this result it can be inferred that in the treatment less fixation of N occurred via biological means. Thus, there was less absorption of S by the dwarf pigeon pea, allowing for greater use of the macronutrient on the part of the corn plants.
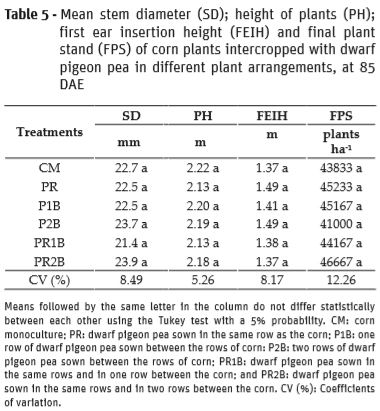
No significant differences were observed for the variables stem diameter (SD), plant height (PH), first ear insertion height (FEIH), final plant stand (FPS) of corn, number of ears per plant (NEP), number of rows of grains in the ear (NRE), number of grains per ear (NGE) and thousand grain weight (TGW) (Tables 5 and 6).
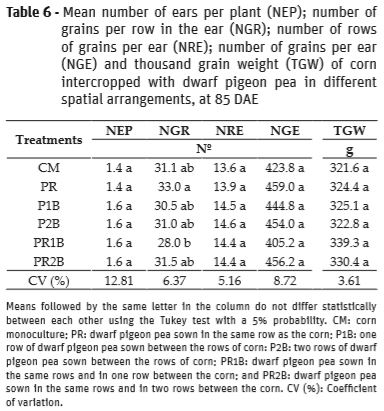
As for the number of grains per row in the ear, there was a significant difference between the arrangements of dwarf pigeon pea (Table 6).
The PR arrangement was superior to the other treatments. According to Balbinot et al. (2005), the number of grains per row directly interferes with ear length and is the yield component that presents the best correlation with productivity, overcoming the other components. Sangoi (2001) points out that the competition of plants for incident solar radiation, nutrients and water determines the ear formation, since it may imply a deficiency of carbon and nitrogen supply to the plants.
For grain yield, it was found that there was a significant difference between the evaluated treatments (Figure 3). The corn intercropped with dwarf pigeon pea sown in the same row as corn and in two rows between them (PR2B) was superior to crop monoculture and to the other plant arrangements, which did not differ from each other. However, all values were lower than those obtained by Arantes et al. (2016), with yield of 9.0 t ha-1, using the same corn variety and intercropping with green manure (Calopogonium mucunoides Desv. (with a density of 21 seeds m-1), Pueraria phaseoloides (Roxb.) Benth (60 seeds m-1) and Neonotonia wightii (Wight & Arn) Lackey) (30 seeds m-1), cultivated under the same environmental conditions.
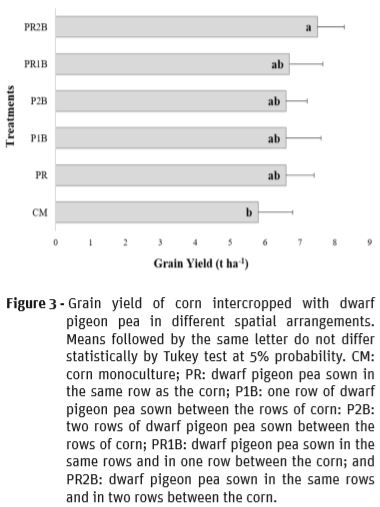
The superiority of PR2B in relation to the other treatments can be attributed to the higher population of dwarf pigeon pea plants in the system, increasing the N uptake through biological fixation, benefiting the nutrient absorption by corn plants. This occurrence can be confirmed by the higher N content observed in corn leaves in the PR2B system (Tables 3 and 4). Spagnollo et al. (2002) also verified a higher corn yield under intercropping compared to corn monoculture, attributing the response to the N supply by green manures (Canavalia ensiformi L., Cajanus cajan L., Stizolobium niveum Kuntze and Glycine sp.).
Corn can be benefited by the N2 fixed by the legume in intercropping, either by the direct excretion of nitrogenous compounds, by the fast decomposition of nodules and roots rich in N (Fustec et al., 2010) or by the direct linking through mycorrhizae in the corn roots and legume roots (Van der Heijden and Horton, 2009). In addition to the N supply, the higher number of legume plants may have improved the soil organic matter content and, consequently, the cycling of other nutrients (Zaccheo et al., 2016), also influencing moisture and weed control (Malik et al., 2006), thus increasing corn grain yield.
CONCLUSIONS
The arrangements of dwarf pigeon pea plants intercropped with corn are viable for the production of higher corn yield. In general, they did not negatively influence the agronomic characteristics and growth of the Poaceae, which shows that regardless of the arrangement of plants the dwarf pigeon pea did not compete with the corn for production factors, such as light, water and nutrients.
The arrangements of dwarf pigeon pea in intercropping did not significantly affect the absorption and accumulation of the macronutrients P, Mg and S by corn, but the arrangement with the greatest population of pigeon pea favored the accumulation of N in corn.
References
Alvarenga, R.C.; Cobucci, T.; Kluthcouski, J.;Wruck, F.J.; Cruz, J.C. & Gontijo Neto M.M. (2006) – A cultura do milho na integração lavoura-pecuária. Informe Agropecuário, vol. 27, p. 106-126. [ Links ]
Arantes, A.C.C.; Fontanetti, A.; Soares, M.R.; Silva Neto, F.J. & Próspero, A.G. (2016) - Agronomic characteristics and yield of organic maize straw intercropped with perennial green manures. Pesquisa Agropecuaria Tropical, vol. 46, n. 3, p. 222-229. http://dx.doi.org/10.1590/1983-40632016v4641054 [ Links ]
Balbinot, A.A.J.; Backes, R.L.; Alves, A.C.; Ogliari, J.B. & Fonseca, J.A. (2005) - Contribuição de componentes de rendimento na produtividade de grãos em variedades de polinização aberta de milho. Revista Brasileira de Agrociência, vol. 11, n. 2, p. 161-166. [ Links ]
Belel, M.D.; Halim, R.A.; Rafii, M.Y. &Saud, H.M. (2014) - Intercropping of corn with some selected legumes for improved forage production: a review. Journal of Agricultural Science, vol. 6, n. 3, p. 48-62. http://dx.doi.org/10.5539/jas.v6n3p48 [ Links ]
Benicasa, P.;Tosti, G.;Tei, F. &Guiducci, M. (2010) - Actual N availability from winter catch crops used for green manuring in maize cultivation. Journal of Sustainable Agriculture, vol. 34, n. 7, p. 705-723. https://doi.org/10.1080/10440046.2010.507452 [ Links ]
Boer, C.A.; Assis, R.L.; Silva, G.P.; Braz, A.J.B.P.; Barroso, A.L.L.; Filho, A.C. & Pires, F.R. (2008) - Biomassa, decomposição e cobertura do solo ocasionada por resíduos culturais de três espécies vegetais na região Centro Oeste do Brasil. Revista Brasileira de Ciência do Solo, vol. 32, n. 2, p. 843-851. http://dx.doi.org/10.1590/S0100-06832008000200038 [ Links ]
Cantarella, H.; Raij, B.V. & Camargo, C.E.O. (1996) - Cereais. In: Raij, B.V.; Cantarella, H.; Quaggio, J.A. & Furlani, M.C. (Eds.) - Recomendações de adubação e calagem para o Estado de São Paulo. Campinas, Instituto Agronômico, p. 45-47. [ Links ]
Casper, B.B.; Cahill Jr., J.F. & Hyatt, L.A. (1998) - Aboveground competition does not alter biomass allocated to roots in Abutilon theophrasti. New Phytologist, vol. 140, n. 2, p. 231-238. http://dx.doi.org/10.1046/j.1469-8137.1998.00271.x [ Links ]
Cheng, Z.H. & Xu, P. (2013) - Lily (Lilium spp.) root exudates exhibit different allelopathies on four vegetable crops. Acta Agriculturae Scandinavica B, vol. 63, n. 2, p. 169-175. https://doi.org/10.1080/09064710.2012.734323 [ Links ]
Cortez, J.W.; Furlani, C.E.O. & Silva, R.P. (2009) - Sistemas de adubação e consórcio de culturas intercalares e seus efeitos nas variáveis de colheita da cultura do milho. Engenharia Agrícola, vol. 29, n. 2, p. 277-287. http://dx.doi.org/10.1590/S0100-69162009000200011 [ Links ]
Cury, J.P.; Santos, J.B.; Silva, E.B.; Byrro, E.C.M.; Braga, R.R.; Carvalho, F.P. & Silva, D.V. (2012) - Acúmulo e partição de nutrientes de cultivares de milho em competição com plantas daninhas. Planta Daninha, vol. 30, n. 2, p. 287-296. http://dx.doi.org/10.1590/S0100-83582012000200007 [ Links ]
Damián-Nava, A.; González-Hernández, V.A.; García, P.S.; Valdívia, C.B.P. &Munõz, M.L. (2006) - Dinámica y diagnóstico nutrimental del guayabo em Iguala, Guerrero, México. Terra Latinoamericana, vol. 24, n. 1, p. 125-132. [ Links ]
Fávero, C.; Jucksch, I.; Alvarenga, R.C. & Costa, L.M. (2001) - Modificações na população de plantas espontâneas na presença de adubos verdes. Pesquisa Agroecuária Brasileira, vol. 36, n. 11, p. 1355-1362. http://dx.doi.org/10.1590/S0100-204X2001001100005 [ Links ]
Fustec, J.; Lesuffleur, F.; Mahieu, S. & Cliquet, J.B. (2010) - Nitrogen rhizodeposition of legumes: a review. Agronomy for Sustainable Development, vol. 30, n. 1, p. 57-66. https://doi.org/10.1051/agro/2009003 [ Links ]
Gitti, D.C.; Arf, O.; Vilela, R.G.; Portugal, J.R.; Kaneko, F.H. & Rodrigues, R.A.F. (2012) - Épocas de semeadura de crotalária em consórcio com milho. Revista Brasileira de Milho e Sorgo, vol. 11, n. 2, p. 156-158. http://dx.doi.org/10.18512/1980-6477/rbms.v11n2p156-168 [ Links ]
Godoy, R.; Bacchi, O.O.S.; Moreira, F.A. & Reichardt, K. (2009) - Evaluation of pigeon pea lines for biological soil decompaction. International Journal of Agronomy, vol. 2009, art. 609717. http://dx.doi.org/10.1155/2009/609717 [ Links ]
Lauer, J.G.; Roth, G.W. & Bertran, M.G. (2004) - Impact of defoliation on corn forage yield. Agronomy Journal, vol. 96, n. 5, p. 459-1463. [ Links ]
Li, Q.Z.; Sun, J.H.; Wei, X.J.; Christie, P.; Zhang, F.S. & Li, L. (2011) - Overyielding and interspecific interactions mediated by nitrogen fertilization in strip intercropping of maize with faba bean, wheat and barley. Plant and Soil, vol. 339, n. 1-2, p. 147–161. https://doi.org/10.1007/s11104-010-0561-5 [ Links ]
Lithourgidis, A.S.; Dordas, C.A.; Damalas, C.A. & Vlachostergios, D.N. (2011) - Annual intercrops: an alternative pathway for sustainable agriculture. Australian Journal of Crop Science, vol. 5, n. 4, p 396-410. [ Links ]
Liu, T.D. & Song, F.B. (2012) - Maize photosynthesis and microclimate within the canopies at grain-filling stage in response to narrow-wide row planting patterns. Photosynthetica, vol. 50, n. 2, p. 215-222. https://doi.org/10.1007/s11099-012-0011-0 [ Links ]
Malavolta, E.; Vitti, G.C. & Oliveira, A.S. (1989) - Avaliação do estado nutricional de plantas: princípios e aplicações. Piracicaba, Potafos, 201 p. [ Links ]
Malik, M.A.; Zahoor, F.; Abbas, S.H. & Ansar, M. (2006) - Comparative study of different herbicides for control of weeds in rainfed maize (Zea mays L.). Islamabad, Weed Science Society of Pakistan, 62 p. [ Links ]
Medeiros, J.C.; Albuquerque, J.A.; Mafra, J.L.; Rosa, J.B. & Gatiboni, L.C. (2008) - Relação cálcio: magnésio do corretivo da acidez do solo na nutrição e no desenvolvimento inicial de plantas de milho em um Cambissolo Húmico Álico. Semina: Ciências Agrárias, vol. 29, n. 4, p. 799-806. [ Links ]
Montañés, L.; Heras, L.; Abadía, J. & Sanz, M. (1993) - Plant analysis interpretation based on a new index: Deviation from optimum percentage (DOP). Journal of Plant Nutrition, vol. 16, n. 7, p. 1289-1308. https://doi.org/10.1080/01904169309364613 [ Links ]
Mushagalusa, G.N.; Ledent, J.F. & Draye, X. (2008) - Shoot and root competition in potato/maize intercropping: Effects on growth and yield. Environmental and Experimental Botany, vol. 64, n. 2, p. 180-188. https://doi.org/10.1016/j.envexpbot.2008.05.008 [ Links ]
Nascimento, F.M.; Bicudo, S.J.; Fernandes, D.M.; Rodrigues, J.G.L. & Fernandes, J.C. (2012) - Diagnose foliar em plantas de milho em sistema de semeadura direta em função de doses e épocas de aplicação de nitrogênio. Applied Research & Agrotechnology, vol. 5, n. 1, p. 67-86. https://doi.org/10.5777/paet.v5i1.1642 [ Links ]
Oliveira, P.; Kluthcouski, J.; Favarinm, J.L. & Santos, D.C. (2011) - Consórcio de milho com braquiária e guandu-anão em sistema de dessecação parcial. Pesquisa Agropecuária Brasileira, vol. 46, n. 10, p. 1184-1192. [ Links ]
Paiva, P.J.R. & Nicodemo, M.L.F. (1993) - Enxofre no sistema solo-planta-animal. Campo Grande, Embrapa, 45 p. [ Links ]
Pariz, C.M.; Andreotti, M.; Azenha, M.V.; Bergamaschine, A.F.; Mello, L.M.M. & Lima, R.C. (2011) - Produtividade de grãos de milho e massa seca de braquiárias em consórcio no sistema de integração lavoura-pecuária. Ciência Rural, vol. 41, n. 5, p. 875-882. [ Links ]
Peng, X.; Zhang, Y.; Cai, J. & Zhang, S. (2009) - Photosynthesis, growth and yield of soybean and maize in a tree-based agroforestry intercropping system on the Loess Plateau. Agroforestry Systems, vol. 76, n. 3, p. 569-577. https://doi.org/10.1007/s10457-009-9227-9 [ Links ]
Ritchie, S.W.; Hanway, J.J. & Benson, G.O. (2003) - Como a planta de milho se desenvolve. Piracicaba, Potafos, 20 p. [ Links ]
Sangoi, L. (2001) - Understanding plant density effects on maize growth and development: an important issue to maximize grain yield. Ciência Rural, vol. 31, n. 1, p. 159-168. http://dx.doi.org/10.1590/S0103-84782001000100027 [ Links ]
Sangoi, L.; Almeida, M.L.; Horn,D.; Bianchet, P.;Gracietti, M.A.; Schmitt, A. & Schweitzer, C. (2004) - Tamanho de semente, profundidade de semeadura e crescimento inicial do milho em duas épocas de semeadura. Revista Brasileira de Milho e Sorgo,vol. 3, n. 3, p. 370-380. http://dx.doi.org/10.18512/1980-6477/rbms.v3n03p%25p [ Links ]
Santos, P.M.; Santos, A.C.; Negreiros Neto, J.V.; Araújo, A.S. & Silva, J.E.C. (2011) - Caracterização de pastagens de capim tanzânia e mombaça consorciados com estilosantes em ecótono de transição Cerrado:Floresta Amazônica. Revista Brasileira de Ciências Agrárias, vol. 6, n. 1, p. 163-173. http://dx.doi.org/10.5039/agraria.v6i1a817 [ Links ]
Spagnollo, E.; Bayer, C.;Wildner, L.P.; Ernani, P.R.; Albuquerque, J.A. & Proença, M.M. (2002) - Leguminosas estivais intercalares como fonte de nitrogênio para o milho, no Sul do Brasil. Revista Brasileira de Ciência do Solo, vol. 26, n. 2, p. 417-423. http://dx.doi.org/10.1590/S0100-06832002000200015 [ Links ]
Van Der Heijden, M.G.A. & Horton, T.R. (2009) - Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. Journal of Ecology, vol. 97, n. 6, p. 1139-1150. http://dx.doi.org/10.1111/j.1365-2745.2009.01570.x [ Links ]
Viera, M.; Schumacher, M.V.; Caldeira, M.V.W. & Watzlawick, L.F. (2013) - Teores de nutrientes em povoamentos monoespecíficos e mistos de Eucalyptus urograndis e Acacia mearnsii em sistema agrossilvicultural. Ciência Florestal, vol. 23, n. 1, p. 67-76. http://dx.doi.org/10.5902/198050988440 [ Links ]
Wu, K.X. & Wu, B.Z. (2014) - Potential environmental benefits of intercropping annual with leguminous perennial crops in Chinese agriculture. Agriculture, Ecosystems and Environment, vol. 188, p. 147-149. https://doi.org/10.1016/j.agee.2014.02.026 [ Links ]
Yusuf, A.A.; Abaidoo, R.C.; Iwuafor, E.N.O.; Olufajo, O.O. & Sanginga, N. (2009) - Rotation effects of grain legumes and fallow on maize yield, microbial biomass and chemical properties of an Alfisol in the Nigerian savanna. Agriculture, Ecosystems & Environment, vol. 129, n. 1-3, p. 325–331. https://doi.org/10.1016/j.agee.2008.10.007 [ Links ]
Zaccheo, P.V.C.; Neves, C.S.V.J.; Mariano, D.C.; Zornenone, T.O.; Higashibara, L.R.; Piccinin, G.G. & Okumura, R.S. (2016) - Green manure in fruticulture: Aspects on soil quality and use in agriculture. African Journal of Agricultural Research, vol. 11, n. 17, p. 1469-1474. https://doi.org/10.5897/AJAR2015.10416 [ Links ]
Zhang, L.; Spiertz, J.H.J.; Zhang, S.; Li, B. & Werf, W. (2007) - Nitrogen economy in relay intercropping systems of wheat and cotton. Plant and Soil, vol. 303, n. 1-2, p. 55-68. https://doi.org/10.1007/s11104-007-9442-y [ Links ]
Zhang, Y.; Liu, J.; Zhang, J.; Liu, H.; Liu, S.; Zhai, L.; Wang, H.; Lei, Q.; Ren, T. & Yin, C. (2015) - Row ratios of intercropping maize and soybean can affect agronomic efficiency of the system and subsequent wheat. PLoS One, vol. 10, n. 6, art. e0129245. https://doi.org/10.1371/journal.pone.0129245 [ Links ]
Received/recebido: 2017.11.20
Received in revised form/recebido em versão revista: 2018.01.30
Aceite/accepted: 2018.02.03

