










Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Ciências Agrárias
Print version ISSN 0871-018X
Rev. de Ciências Agrárias vol.39 no.1 Lisboa Mar. 2016
https://doi.org/10.19084/RCA15111
ARTIGO
Os compostos no controlo de doenças das plantas
Control of plant diseases with composts
Mário Reis*
*Departamento de Ciencias Biológicas e Bioengenharia, Faculdade de Ciências e Tecnologia, Universidade do Algarve, Campus de Gambelas, Edifício 8, 8005-139 Faro, Portugal. E-mail: mreis@ualg.pt
RESUMO
A compostagem permite obter produtos, designados compostados ou compostos, que em alguns casos, apresentam a capacidade de reduzir ou eliminar doenças das plantas com origem no solo – capacidade supressiva ou supressividade – o que lhes confere um interesse acrescido ao de fertilizantes agrícolas, objectivo com que habitualmente se empregam. Neste trabalho, após breve descrição da compostagem, são referidos os mecanismos de supressividade, as condições para a obtenção de compostos supressivos, os métodos de quantificação da supressividade e referem-se alguns exemplos de compostos com capacidade supressiva. O conhecimento sobre a capacidade supressiva dos compostos é uma área de estudo com grande carência de conhecimento, mas que apresenta um enorme interesse, em particular numa perspectiva de produção agrícola mais sustentável e com menor impacte ambiental.
Palavras-chave: antagonistas, compostagem, composto, controlo biológico
ABSTRACT
Composts, obtained through the biological process of composting, frequently show the capacity to reduce or eliminate plant diseases – supressivity or supressive capacity- that increases the interest for its use in agriculture. After a short characterisation of composting, the mechanisms of suppressivity are discussed, as well as the conditions to obtain supressive composts and the methods to determine and quantify the suppressive capacity. Some supressive composts, used as substrate components or soil amendments, are mentioned. More research on supressivity is needed, due to the high potential for the use of composts towards a more environmental friendly and sustainable agriculture.
Keywords: antagonists, biological control, compost, composting
Introdução
A compostagem é um processo biológico de transformação de resíduos orgânicos pelo qual se obtêm produtos, designados compostados ou compostos, que podem ser empregues como correctivos orgânicos, fertilizantes ou componentes de substratos hortícolas. A aplicação de compostos ao solo ou na formulação de substratos hortícolas, pode reduzir a ocorrência de algumas doenças das plantas com origem no solo, bem como de doenças da parte aérea. Contudo, ocorrem por vezes resultados ambíguos ou contraditórios, em consequência da diversidade de condições de obtenção e emprego dos compostos, e da complexidade dos processos de controlo dos microrganismos fitopatogénicos.
Os compostos: obtenção e características
Composto é um produto estabilizado e higienizado, benéfico para o crescimento das plantas, resultante de compostagem (Zucconi e Bertoldi, 1987). A compostagem é um processo biológico controlado, no qual microrganismos atacam a matéria orgânica, obtendo energia e nutrientes para a sua actividade, sendo o produto final – o composto ou compostado - constituído pelas fracções mais resistentes da matéria orgânica, pelos produtos da sua decomposição, por produtos sintetizados pelos microrganismos durante o processo, e ainda pelos organismos, vivos e mortos. Tipicamente, a compostagem desenrola-se em três fases: a inicial ou mesofílica, que dura normalmente poucos dias, durante a qual a temperatura sobe a 40 - 50ºC, o pH baixa e são metabolizados os componentes mais biodegradáveis da matéria orgânica; a intermédia ou termofilica, que dura de poucas semanas a alguns meses, a temperatura atinge 70ºC ou mais, o pH sobe, os microorganismos termofílicos degradam a matéria orgânica até esgotar a fracção mais facilmente degradável; e a fase de estabilização ou maturação, durante a qual se reduz a actividade microbiana, a temperatura diminui até à temperatura ambiente e os microrganismos mesofílicos recolonizam o meio (Zucconi e Bertoldi, 1987).
Durante a compostagem aumenta a estabilidade física e química da matéria orgânica remanescente; são inviabilizadas sementes de infestantes e patogenos, e criadas condições desfavoráveis à actividade de outros; reduz-se a quantidade de produtos químicos indesejáveis e melhora a facilidade de manuseamento do material (Daft et al., 1979; Spencer e Benson, 1982). A qualidade do composto é quantificável através de variáveis físicas, químicas e biológicas, e de índices específicos (Inbar et al., 1993).
O conceito de supressividade
Solos supressivos e solos conducivos
Baker e Cook (1974) definiram solos supressivos como aqueles onde um patogeno não se consegue estabelecer ou persistir; consegue-se estabelecer mas causa poucos ou nenhuns danos; ou estabelece-se e causa danos, mas com importância descrescente, apesar de se manter no solo. Ferraz (1992) considera supressivos, os solos capazes de reduzir ou eliminar doenças com origem no solo, mesmo na presença de elevada quantidade de inóculo. Em oposição, os solos onde pouco inóculo é suficiente para a manifestação da doença designam-se solos conducivos ou permissivos (Borges e Jorge-Silva, 1992), existindo uma gama de comportamentos intermédios. A capacidade supressiva dos solos foi assinalada há mais de 100 anos (Ferraz, 1992), mas só mais tarde se associou o controlo das doenças do solo à presença de outros microrganismos no solo, e se assumiu ser possível actuar sobre o grau de controlo biológico por modificação das práticas culturais (Sanford, 1926). Assim, embora a capacidade supressiva de um solo esteja associada às suas propriedades (Persson e Olsson, 2000), pode ser melhorada de forma a conseguir-se o controlo biológico das doenças com origem no solo (Alabouvette, 1999), através da infestação do solo com microrganismos específicos – antagonistas - e de práticas culturais tendentes a melhorar a supressividade natural dos solos (Sturz et al., 1997; Bailey e Lazarovits, 2003), tais como como a incorporação de materiais orgânicos no solo (Cook, 1993). Também algumas doenças da parte aérea podem ser controladas pela acção da microflora do solo (Kloepper et al., 1999; Segarra et al., 2009).
Alguns compostos manifestam também acção supressiva quando aplicados no solo ou utilizados como componentes de substratos.
Mecanismos de supressividade
O controlo de doenças com origem no solo, conseguido pela aplicação de compostos, é em parte associado às suas propriedades químicas, como a presença de toxinas alelopáticas, nutrientes (e.g.: azoto, ferro e fósforo), relação carbono/ azoto, pH e salinidade. Com a mineralização progressiva dos compostos, a importância destes inibidores químicos parece decrescer, aumentando o papel dos microrganismos na capacidade supressiva exercida pelo composto (Lootsma e Scholte, 1997), os quais obtêm a energia que necessitam da matéria orgânica do composto, da matéria orgânica solo e da disponibilizada através dos exsudados radiculares (Shiomi et al., 1999).
Os componentes da tripla interacção: hospedeiro - parasita - população microbiana, são afectados por factores ambientais (Park, 1963), e em particular, no caso dos parasitas da raiz, pela presença e actividade dos microrganismos do solo, onde se destacam bactérias, actinomicetas, protozoários, nemátodos, fungos e oomicetas. Por simplificação, tratamos neste trabalho os oomicetas, incluídos actualmente no reino Chromista (e.g.: géneros Phytophthora, Olpidium, Plasmopara), como fungos (reino Fungi) (Ferreira, 2008). Os fungos assumem importância particular devido ao elevado número de doenças que podem causar nas plantas. Do ponto de vista ecológico, Garrett (1956) classificou os fungos em dois grandes grupos: os habitantes do solo e os habitantes das raízes, ambos com espécies com capacidade de provocar doenças nas plantas (Figura 1).
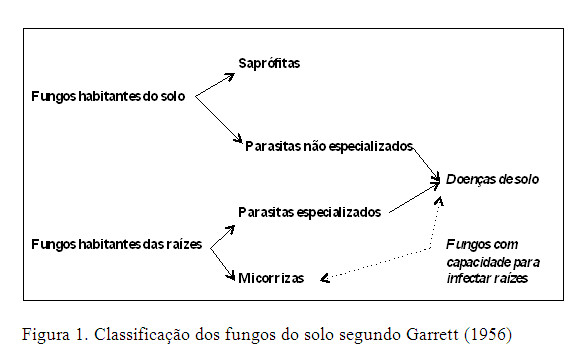
Os fungos habitantes do solo têm uma distribuição generalizada nos solos, onde podem viver indefinidamente como saprófitas (e.g.: Aspergillus spp., Penicillium spp., Trichoderma spp.), ou ter uma existência parasítica (e.g.: Rizoctonia solani, Fusarium culmorum, Pythium spp., Phytophthora spp.), infectando raízes e causando doenças em inúmeros hospedeiros, fazendo com que as suas populações se mantenham relativamente estáveis, independentemente da presença do hospedeiro (Ferraz, 1985).
Pelo contrário, os fungos habitantes das raízes apresentam uma fase parasítica, em expansão, na presença do hospedeiro, e uma fase saprofítica, em declínio, nos tecidos mortos do hospedeiro. Entre os fungos habitantes das raízes, incluem-se espécies com fraca ou nenhuma capacidade de sobrevivência como saprófitas na ausência do hospedeiro, embora sobrevivam no solo sob a forma de estruturas de resistência (e.g.: F. oxysporum, Verticillium spp.). Os fungos parasitas das raízes são mais sensíveis aos antibióticos produzidos pelos microrganismos do solo, só se desenvolvem no solo a partir de um substrato, invadem somente tecidos vivos ou senescentes e possuem uma distribuição localizada devido à sua associação com o hospedeiro. Além destas características biológicas, outras, como o seu inoculo potencial e a capacidade de sobrevivência, são determinantes para a sua capacidade de provocar doenças nas plantas (Ferraz, 1985).
A influência dos microrganismos do solo sobre a actividade e sobrevivência dos microrganismos patogénicos pode-se exercer através de: competição por nutrientes, oxigénio ou pontos de invasão do hospedeiro; antibiose; microbiostase; produção de compostos capazes de anular substâncias tóxicas; hiperparasitismo; predação; hipovirulência e pelo estímulo de mecanismos de resistência do hospedeiro (Garrett, 1970; Maurhofer et al., 1994; Lugtenberg e Kamilova, 2009).
A interacção entre o meio e os microrganismos, e entre microrganismos, permite distinguir dois tipos de supressividade: a geral e a específica (Baker e Cook, 1974). A supressividade geral resulta da actividade de diferentes espécies de microrganismos que, ao competirem, limitam a disponibilidade de recursos para os patogenos. Sendo os microrganismos fitopatogénicos do solo caracterizados normalmente por uma reduzida capacidade de competição saprofítica (excepto os fungos habitantes do solo não especializados), a sua actividade é determinada pela microflora do solo. Os propágulos dos patogenos controlados pelo mecanismo de supressividade geral não se reduzem rapidamente num meio supressivo, são pequenos (menos de 200 mm de diâmetro), não armazenam muitos nutrientes e dependem da disponibilidade de fontes de carbono para a germinação e infecção (Chen et al., 1988a,b; Nelson, 1990), como por exemplo Pythium e Phytopthora spp. (Hardy e Sivasithamparam, 1991a,b; Boehm et al., 1993). A eficácia dos microrganismos no controlo biológico dos patogéneos, dependendo da disponibilidade de energia na matéria orgânica, pode ser a razão para a variabilidade observada na eficácia do controlo de Pythium (Boehm et al., 1993).
Na supressividade específica o controlo dos patogenos resulta da acção de um ou de poucos organismos – designados antagonistas - como sucede relativamente a R. solani e Sclerotium rolfsii, que podem ser parasitados por hiperparasitas específicos como Gliocladium virens (Lewis et al., 1995), espécies binucleadas de Rhizoctonia (Villajuanabgona et al., 1996) ou Pseudomonas fluorescens (Couillerot et al., 2009). A inibição de Fusarium oxysporum f. sp. basilica é conseguida por Glomus mosseae (Toussaint et al., 2008). F. oxysporum f. sp. dianthi e F. o. f. sp. lycopersici são inibidos por Trichoderma asperellum (Cotxarrera et al., 2002; Sant et al., 2010), embora a germinação dos clamidósporos de F. o. f. sp. lycopersici seja relacionada também com o pH do meio (Shiau et al., 1994). A produção, por alguns microrganismos, de antibióticos (Hill et al., 1994; Pal e McSpadden Gardener, 2006; Zaccardelli et al., 2013) ou enzimas (Punja e Zhang, 1993; McSpadden Gardener et al., 2005) são outros processos de supressividade específica, bem como a indução de resistência sistémica (Larkin et al., 1996). A acção supressiva de um microrganismo pode-se exercer com maior eficácia pela combinação de várias daquelas acções (Duijff et al., 1993).
Relacionando a classificação dos fungos do solo de Garrett (1956) e os tipos de supressividade enunciados por Baker e Cook (1974), observa-se que os fungos habitantes do solo são controlados segundo o mecanismo de supressividade geral (Figura 2).
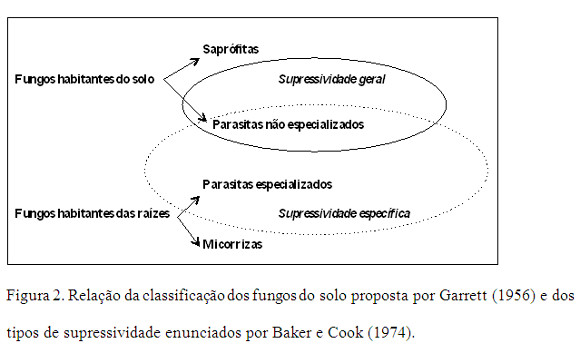
Sendo os fungos habitantes do solo de ocorrência frequente e com elevada capacidade saprofítica (e.g.: Pythium spp., Phytophthora spp.), são controlados através da competição imposta pelas activas e diversificadas populações microbianas introduzidas pela aplicação de composto estabilizado, onde existem fontes alimentares limitadas. Contudo, alguns dos fungos habitantes do solo - os não especializados - que apresentam volumosas estruturas de resistência (e.g.: R. solani e S. rolsfii) só são controlados pelo mecanismo de supressividade específica, tal como os fungos habitantes das raízes (e.g.: F. oxysporum e Verticillium spp.) (Ferraz, 1985).
Em síntese, a supressividade geral está relacionada com a biomassa total de microganismos do solo, e não apenas com uma única espécie, que competem com o patogeno, e é estimulada por acções que promovam a actividade microbiológica no solo, como a adição de materia orgânica ou certas práticas culturais. A supressividade especifica caracteriza-se pela sua fácil transferência, pois os responsáveis, pelo menos em parte, são espécies ou grupos determinados de microganismos. Contudo, os solos supressivos devem normamente esta característica à combinação dos processos de supressividade geral e específica (Weller et al., 2002).
Os compostos como agentes de supressividade
Condições para a obtenção de compostos supressivos
Desde que, em meados do séc. XX, se observou capacidade supressiva em compostos (Howard, 1945), vários destes produtos, principalmente compostos de casca de árvores, têm sido usados com sucesso no controlo de doenças com origem no solo (Hoitink, 2001; Bailey e Lazarovits, 2003). A capacidade supressiva manifesta-se nos compostos (Nelson e Hoitink, 1983; Chen e Hadar, 1987), embora por vezes a compostagem não aumente essa capacidade (Voland e Epstein, 1994) ou tenha eliminado as causas inibidoras dos microrganismos fitopatogénicos (Nelson e Hoitink, 1982). Na prática, a supressividade induzida pelos compostos depende de vários factores, nomeadamente: a interacção agente patogénico - hospedeiro - microrganismos (na rizosfera e no composto), e as características físicas e químicas e disponibilidade de nutrientes do meio (Hoitink et al., 1993).
Hoitink et al. (1993) indica como condições para o sucesso no uso de compostos no controlo biológico de doenças do solo: (i) a morte dos organismos patogénicos durante a compostagem, (ii) a presença de uma activa população de microrganismos benéficos no composto, (iii) a estabilização adequada da matéria orgânica e (iv) as características físicas e químicas do composto.
Morte dos organismos patogénicos durante a compostagem
A maioria dos patogenos são sensíveis às condições ambientais durante a compostagem, podendo os compostos ser utilizados com segurança, à excepção dos obtidos com material infectado com alguns vírus, como TMV e TNV. A inactivação dos patogenos na compostagem – o que pode explicar a não manifestação de doença ao empregar os compostos (Hoitink et al., 1975) - resulta da elevação da temperatura, da acção de produtos tóxicos libertados durante a decomposição da matéria orgânica (e.g.: etanol, formaldeído e amoníaco) e de processos de antagonismo, actuando sucessivamente ou em simultâneo (Berestetsky e Kravchenko, 1984; Bollen, 1993).
A resistência dos patogenos à temperatura elevada durante a compostagem depende de factores como a densidade da população e a humidade do material. Poucos fungos permanecem activos quando a temperatura ultrapassa 70 oC (Hartmann et al., 1990), estando a inactivação dos mais resistentes associada à humidade (Bollen, 1993) e ao pH do material (Ylimäki et al., 1983).
Embora muitas espécies de fungos sejam eliminadas durante a compostagem, algumas podem resistir (Bollen, 1993). No caso dos vírus, mesmo que estes não sejam destruídos mas apenas os seus vectores, isto pode ser o suficiente para a utilização segura dos compostos, se no local em que o composto vai ser utilizado os vectores não existirem. Se os vectores dos vírus forem nemátodos, a infecção do composto não ocorre pois as espécies transmissoras de vírus alimentam-se apenas a partir de plantas vivas e não do composto; se forem fungos produtores de zoósporos, a possibilidade de transmissão depende do processo de transmissão, por exemplo, é possível com fungos do género Olpidium e pouco provável nos do género Polymyxia (Bollen, 1993). Os nemátodos são bastante sensíveis à temperatura, sendo destruídos a temperatura abaixo de 50 oC (Hartmann e Kester, 1990). A maioria das bactérias não sobrevive às condições normais da compostagem, excepto as bactérias produtoras de esporos, cuja destruição exige temperatura superior a 70 oC (Hartmann e Kester, 1990).
Presença de uma activa população microbiana de organismos benéficos
Competição e antagonismo são processos importantes para o controlo dos patogenos durante a compostagem (Bertoldi et al., 1991), embora alguns autores atribuam ao antagonismo um papel pouco relevante (Bollen, 1993). Contudo, a exposição a alta temperatura e a produtos tóxicos pode enfraquecer os patogenos, tornando-os mais sensíveis ao antagonismo que se estabelece após a fase termofilica da compostagem (Yuen e Raabe, 1984). A indução de um nível consistente de supressividade no composto implica a recolonização por organismos mesófilos após a fase termofílica. No final da compostagem, procura-se a recolonização natural do meio pela maior diversidade possível de mesófilos, ou combinar este objectivo com a introdução de microrganismos antagonistas seleccionados (Hoitink, 1987; Sivapalan et al., 1994). No entanto, presença de microrganismos benéficos não significa eficácia, podendo uma menor população ser mais eficaz nesse controlo que outra maior (Nelson e Hoitink, 1983; Nelson et al., 1983).
Se a capacidade supressiva de um composto tiver sido eliminada por um tratamento térmico, pode ser restaurada através da mistura de uma pequena quantidade de composto não tratado termicamente (Westphal e Becker, 2000) ou através da infestação com microrganismos específicos, como Trichoderma hamatum e Flavobacterium balustinum (Trillas-Gay et al., 1986). A manutenção do efeito supressivo é apenas possível nos meios com suficiente capacidade de suporte da actividade microbiológica e reservas de polissacáridos (Chen et al., 1988a, b; Hoitink et al., 1993), o que pode ocorrer não apenas num solo mas também mesmo no cultivo sem solo, por exemplo em lã-de-rocha (Tu et al., 1999).
Estabilização adequada da matéria orgânica
O grau de decomposição da matéria orgânica determina a actividade da microflora benéfica e o inoculo potencial dos patogenos. A matéria orgânica fresca pode servir de fonte alimentar aos patogenos e aumentar o seu inoculo potencial (Garrett, 1962; Hoitink, et al., 1993).
Um aspecto muito importante na redução de bactérias e fungos patogénicos em compostos, é o facto de estes microrganismos normalmente apenas conseguirem utilizar produtos facilmente assimiláveis (e.g.: álcoois simples, ácidos orgânicos ou açúcares), não se multiplicando em meios apenas com celulose, lenhina ou substâncias húmicas. Após a compostagem, quando o compostado se encontra enriquecido em celulose, lenhina e substancias húmicas, e pobre em matéria orgânica facilmente degradável, as bactérias e fungos fitopatogénicos ficam em desvantagem perante os microrganismos saprófitas.
Além disso, a população total de microrganismos é numerosa relativamente à dos patogenos, gerando um elevado grau de antagonismo, o que associado à grande competição trófica coloca os patogenos em desvantagem (Bertoldi et al., 1991).
Características físicas e químicas do composto empregue
As características dos compostos afectam a susceptibilidade das plantas aos patogenos, e a actividade destes e dos antagonistas (Score e Palfreyman, 1994) observando-se, por exemplo, diferenças de capacidade supressiva, no solo e in vitro, em consequência da variação do pH do meio (Trillas-Gay et al., 1986; Shiau et al., 1994; Borrero et al., 2004).
Os compostos que libertam muito azoto contribuem para o aumento da manifestação de fusarioses (Hoitink et al., 1997), enquanto que a sua deficiência em azoto as pode reduzir, desde que o composto esteja adequadamente colonizado (Trillas-Gay et al., 1986). Compostos com salinidade elevada acentuam a manifestação de Pythium e de Phytophthora (Hoitink et al., 2001). A aplicação de doses elevadas de fertilizantes ao composto, particularmente azoto e fósforo, pode controlar o desenvolvimento de Phoma chrysanthemicola em crisântemo (Menzies e Colhoum, 1976). O potencial matrico afecta o desenvolvimento microbiano, nomeadamente a formação de esporângios e a libertação de zoósporos e outras estruturas reprodutivas (Spencer e Benson, 1982). Um baixo potencial de água estimula o crescimento de muitos patógenos (Lootsma e Scholte, 1997), mas noutros casos, um maior teor de humidade, permite o desenvolvimento de uma maior população de bactérias antagonistas (Clulow et al., 1995).
Em 17 compostos de vários resíduos agro-industriais, Suárez-Estrella et al. (2012) observaram maior efeito supressivo quando o pH era próximo do neutro, a condutividade eléctrica inferior a 3 dS m-1 e a matéria orgânica superior a 85%
Previsão e quantificação da capacidade supressiva
De um modo previsível, é possível usar práticas de controlo biológico recorrendo a compostos, particularmente no cultivo sem solo em substratos (Hoitink et al., 1993). Contudo, é indispensável um conhecimento preciso das características dos compostos, nomeadamente do grau de estabilidade da matéria orgânica, salinidade e disponibilidade de azoto, sob pena de se obterem resultados muito diversos.
Para quantificar o efeito de supressividade geral utilizam-se técnicas que permitem determinar a absorção de nutrientes pelos microrganismos do solo, a sua actividade ou a sua biomassa (Chen et al., 1988b). Estas determinações permitiram obter métodos para testar a supressividade natural (Chen et al., 1988a; Inbar et al., 1991) e a criação de directivas para a formulação de substratos supressivos (Chen et al., 1988b). Relacionando o conhecimento sobre as substâncias absorvidas pelos microrganismos com o das populações de determinados microrganismos, pode-se determinar o potencial supressivo de um composto.
Compostos com capacidade supressiva
A partir dos anos 60 do séc. XX utilizaram-se com êxito compostos de casca de árvores na preparação de substratos hortícolas, obtendo-se melhores resultados de cultivo que os conseguidos, até aí, com as tradicionais misturas à base de turfa, esterilizada ou não (Daft et al., 1979), registando-se em alguns casos, uma menor incidência de doenças de solo (Hoitink et al., 1975; Nelson e Hoitink, 1982). No entanto, a origem da casca e as condições de compostagem determinam a capacidade supressiva do composto, pelo que esta pode variar com o lote do produto (Daft et al., 1979), o que justifica o elevado rigor na preparação dos compostos para conseguir manter uma aceitável homogeneidade dos produtos.
Outros resíduos têm sido empregues com sucesso na preparação de compostos supressivos, destacando-se a casca de algumas espécies de eucalipto (Handreck e Black, 1991), bagaços de uva e de cana-do-açúcar, laranjas de refugo, resíduos da produção de cogumelos e da manutenção dos espaços verdes (Chen e Hadar, 1987; Martinez, 1989; Theodore e Toribio, 1995; Nakasaki et al., 1998; Szczech et al., 1997; Coelho e Reis, 2011; Sua´rez-Estrella et al., 2012), resíduos sólidos urbanos, produtos de vermicultura (Kuter et al., 1988; Szczech, 1999) e resíduos da pecuária (Martinez, 1989).
A utilização de compostos no cultivo das plantas pode, por isso, contribuir para a sustentabilidade da actividade humana, permitindo reciclar resíduos de origem diversa, com benefícios económicos e ambientais, sobretudo pela redução do consumo de outros fertilizantes e de produtos fitossanitários.
Agradecimentos
Ao Professor Doutor José P. F. Ferraz e ao Professor Doutor António A. Monteiro pelo seu apoio na elaboração deste trabalho.
Referências bibliográficas
Alabouvette, C. (1999) - Fusarium wilt suppressive soils: an example of disease-suppressive soils. Australasian Plant Pathology, vol. 28, n. 1, p. 57-64. http://dx.doi.org/10.1071/AP99008 [ Links ]
Bailey, K.L. e Lazarovits, G. (2003) - Soil agroecosystems: impacts of management on soil health and crop diseases. Soil and Tillage Research, vol. 72, n. 2, p. 169–180. http://dx.doi.org/10.1016/S0167-1987(03)00086-2 [ Links ]
Baker, K.F. e Cook, R.J. (1974) - Biological Control of Plant Pathogens. St. Paul, Minnesota, American Phytopathological Society, 433 p. [ Links ]
Berestetsky, O.A. e Kravchenko, L.V. (1984) - Volatile products of plant residue decomposition and their effect on soil microflora. In: J. Szegi (Ed.) - Soil Biology and Conservation of the Biosfere, Vol. 1. Budapeste, Akadémiai Kaidó, p. 419-425. [ Links ]
Bertoldi, M. de; Zucconi, F. e Civilini, M. (1991) - Temperature, pathogen control and product quality. In: The Staff of BioCycle (Eds.) - The Biocycle guide to the Art & Science of Composting. Emmaus, Pensylvania, EUA, The JG Press, Inc., p. 195-198. [ Links ]
Boehm, M.J.; Madden, L.V. e Hoitink, H.A.J. (1993) - Effect of organic matter decomposition level on bacterial species diversity and composition in relationship to Pythium damping-off severity. Applied and Environmental Microbiology, vol. 59, n. 12, p. 4171-4179. [ Links ]
Bollen, G.J. (1993) - Factors Involved in Inactivation of Plant Pathogens During Composting of Crop Residues, In: Hoitink H.A.J. e Keener H.M. (Eds.) - Science and Engineering of Composting: Design, Environmental, Microbiological and Utilization Aspects. Worthington, EUA, Renaissance Publ., p. 301-318. [ Links ]
Borges, M. de L.V. e Jorge-Silva, M.L. (1992) - Solarização do solo e antagonistas de fungos fitopatogénicos. Revista de Ciências Agrárias, vol. 15, n.1-2, p. 281-291. [ Links ]
Borrero, C.; Trillas, M.I.; Ordovás, J.; Tello, J.C. e Aviles, M. (2004) - Predictive factors for the suppression of Fusarium wilt of tomato in plant growth media. Phytopathology, vol. 94, n. 10, p. 1094-1101. http://dx.doi.org/10.1094/PHYTO.2004.94.10.1094 [ Links ]
Chen, Y. e Hadar, Y. (1987) - Composting and use of agricultural wastes in container media, In: Bertoldi, M. de et al. (Eds.) - Compost: Production, Quality and Use. Essex, Reino Unido, Elsevier Appl. Sci. Publ. Ltd., p. 71-77. [ Links ]
Chen, W.D.; Hoitink, H.A.J. e Madden, L.V. (1988a) - Microbial activity and biomass in container medis for predicting suppressiveness to damping-off caused by Pythium ultimum. Phytopathology, vol. 78, n. 3, p. 1447-1450. http://dx.doi.org/10.1094/Phyto-78-1447. [ Links ]
Chen, W.D.; Hoitink, H.A.J.; Schmitthenner, A.F. e Tuovinen, O.H. (1988b) - The role of microbial activity in suppression of damping-off caused by Pythium ultimum. Phytopathology, 78, n.3, p. 314-322. http://dx.doi.org/10.1094/Phyto-78-314 . [ Links ]
Clulow, S.A.; Stewart, H.E.; Dashwood, E.P. e Wastie, R.L. (1995) - Tuber surface microorganisms influence the susceptibility of potato tubers to late blight. Annals of Applied Biology, vol. 126, n. 1, p. 33-43. http://dx.doi.org/10.1111/j.1744-7348.1995.tb05001.x [ Links ]
Coelho, L. e Reis, M. (2011) - Controlling Rhizoctonia solani in cucumber using compost of agro-industrial wastes. In: Livro de resumos do International Symposium on Growing Media, Composting and Substrate Analysis, Barcelona, Espanha, 88. [ Links ]
Cook, R.J. (1993) - Making greater use of introduced microorganisms for biological control of plant pathogens. Annual Review of Phytopathology, vol. 31, p. 53-80. DOI: http://dx.doi.org/10.1146/annurev.py.31.090193.000413 [ Links ]
Cotxarrera, L.; Trillas-Gay, M.I.; Steinberg C. e Alabouvette, C. (2002) - Use of sewage sludge compost and Trichoderma asperellum isolates to suppress Fusarium wilt of tomato. Soil Biology & Biochemistry, vol. 34, n. 4, p. 467-476. http://dx.doi.org/10.1016/S0038-0717(01)00205-X [ Links ]
Couillerot, O.; Prigent-Combaret, C.; Caballero-Mellado, J. e Moënne-Loccoz, Y. (2009) - Pseudomonas fluorescens and closely related fluorescent pseudomonads as biocontrol agents of soil-borne phytopathogens. Letters in Applied Microbiology, vol. 48, p. 505–512. http://dx.doi.org/10.1111/j.1472-765X.2009.02566.x [ Links ]
Daft, G.C.; Poole, H.A. e Hoitink, H.A.J. (1979) - Composted hardwood bark: a substitute for steam sterilization and fungicide drenches for control of Poinsettia crown and root rot. HortScience, vol. 14, p. 185-187. [ Links ]
Duijff, B.J.; Meijer, J.W.; Bakker, P.A.H.M. e Schippers, B. (1993) - Siderophore-mediated competition for iron and induced resistance in the suppression of Fusarium wilt of carnation by fluorescent Pseudomonas spp. Netherlands Journal of Plant Pathology, vol. 99, n. 5-6, p. 277-289. http://dx.doi.org/10.1007/BF01974309 [ Links ]
Ferraz, J.F.P. (1985) - Doenças radiculares causadas por fungos. I - Uma perspectiva ecológica. Curso de Aperfeiçoamento de Patologia das Plantas Tropicais e Subtropicais. Oeiras 14-31 de Outubro. [ Links ]
Ferraz, J.F.P. (1992) - Os microrganismos antagonistas no controlo das doenças radiculares. ASGARVE, Boletim Informativo, vol. 12, p. 5-7. [ Links ]
Ferreira, R.B. (2008) – Fungos patogénicos das plantas. A batalha química para a patogénese. Agros, n. 2, p. 4-13. [ Links ]
Garrett, S.D. (1956) - Biology of Root-Infecting Fungi. NY, Cambr. Univ. Press, 288 p. [ Links ]
Garrett, S.D. (1962) - Decomposition of cellulose in soil by Rizoctonia solani Kuhn. Transactions of the British Mycological Society, vol. 45, p. 114-120. [ Links ]
Garrett, S.D. (1970) - Pathogenic Root-Infecting Fungi. Nova Iorque, Cambridge Univ. Press, 294 p. [ Links ]
Handreck, K. e Black, N. (1991) - Growing media for ornamental plants and turf. 4ª ed. New Sowth Wales Univ. Press, Kensington, NSW Australia, 401 p. [ Links ]
Hardy, G.E. e Sivasithamparam, K. (1991a) - Effects of sterile and non-sterile leachates extracted from composted eucalyptus bark and pine-bark container media on Phytophthora spp. Soil Biology and Biochemistry, vol. 23, n. 1, p. 25-50. http://dx.doi.org/10.1016/0038-0717(91)90158-G [ Links ]
Hardy, G.E. e Sivasithamparam, K. (1991b) - How container media and matric potential affect the production of sporangia, oospores and chlamydospores by three Phytophthora species. Soil Biology and Biochemistry, vol. 23, n. 1, p. 31-39. http://dx.doi.org/10.1016/0038-0717(91)90159-H [ Links ]
Hartmann, H.T.; Kester D.E. e Davies F.T. (1990) - Plant Propagation. Principles and practices. 5ª ed. New Jersey, EUA, Prentice-Hall, Inc., Englewood Cliffs, 647 p. [ Links ]
Hill, D.S.; Stein, J.I.; Torkeewitz, N.R.; Morse, A.M.; Howell, C.R.; Pachalatko, J.P.; Becker, J.O. e Lignon, J.M. (1994) - Cloning of genes involved in the synthesis of pyrrolnitrin from Pseudomonas fluorescens and role of pyrrolnitrin synthesis in biological control of plant disease. Applied and Environmental Microbiology, vol. 60, n. 1, p. 78-85. [ Links ]
Hoitink, H.A.J.; Schmitthenner, A.F. e Herr L.J. (1975) - Composted bark for control of root rot in ornamentals. Ohio Florists Association Bulletin, vol. 60, p. 25-26. [ Links ]
Hoitink, H.A.J. (1987) - Control of cyclamen Fusarium wilt - A preliminary report. Ohio Florists Association Bulletin, n. 693, p. 1-3. [ Links ]
Hoitink, H.A.J.; Boehm, M.J. e Hadar, Y. (1993) - Mechanisms of Suppression of Soilborn Plant Pathogens, In: Hoitink H.A.J. e Keener H.M. (Eds.) - Science and Engineering of Composting: Design, Environmental, Microbiological and Utilization Aspects. Worthington, EUA, Renaissance Publications. p. 601-621. [ Links ]
Hoitink, H.A.J.; Krause, M.S. e Han, D.Y. (2001) - Spectrum and mechanisms of plant disease control with composts. In: Stofella P.J. e Khan B.A. (Eds.) - Compost Utilization in Horticultural Cropping Systems. Boca Raton, Lewis Publi., p. 263-273. [ Links ]
Howard, A. (1945) - Farming and gardening for health or disease. Faber & Faber, 282 p. [ Links ]
Inbar, Y.; Bohem, M.J. e Hoitink, H.A.J. (1991) - Hydrolysis of fluorescein diacetate in sphagnum peat container media for predicying suppressiveness to damping-off caused by Pythium ultimum. Soil Biology and Biochemistry, vol. 23, n. 5, p. 479-483. http://dx.doi.org/10.1016/0038-0717(91)90013-A [ Links ]
Inbar, Y.; Chen, Y. e Hoitink, H.A.J. (1993) - Properties for Establishing Standards for Utilization of Composts. In: Hoitink, H.A.J. e Keener, H.M. (Eds.) - Science and Engineering of Composting: Design, Environmental, Microbiological and Utilization Aspects. Renaissance Publi. Worthington, p. 668-694. [ Links ]
Kloepper, J.W.; Rodriguez-Kabana, R.; Zehnder, G.W.; Murphy, J.F.; Sikora, E. e Fernandez, C. (1999) - Plant root-bacterial interactions in biological control of soilborne diseases and potential extension to systemic and foliar diseases. Australasian Plant Pathology, vol. 28, n. 1, p. 21-26. http://dx.doi.org/10.1071/AP99003 [ Links ]
Recebido/received: 2015.01.30
Recebido em versão revista/received in revised form: 2015.09.11
Aceite/accepted: 2015.09.14

