










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.38 no.3 Lisboa set. 2015
ARTIGO
Otimização da produção de plantas in vitro de cultivares de Ipomoea batatas
Optimization of the in vitro plants production of Ipomoea batatas cultivars
Rejane Flores1,*, Lilian P. Maggio1, Paola Z. Flôres1, Glaucia S. Bempck1, Neiva M.F. Auler1, João F.C. Carvalho1, Rodrigo S. Godoi1, Simone M. Franzin1, Lara Becker1 e Tatiana M. Silveira1
1Instituto Federal de Educação, Ciência e Tecnologia Farroupilha (IF Farroupilha), Campus São Vicente do Sul, Centro de Recursos Naturais, Rua 20 de setembro, s/n, 97420-000 - São Vicente do Sul, RS – Brasil. E-mails: * rejane.flores@yahoo.com.br, author for correspondence; lilianmaggio@yahoo.com.br; paolazuquetto@gmail.com; bempck.glaucia@yahoo.com.br; frizonauler@yahoo.com.br; joão.carvalho@iffarroupilha.edu.br; rodrigo.godoi@iffarroupilha.edu.br; simone.franzin@iffarroupilha.edu.br; lara.becker@iffarroupilha.edu.br; tatiana.silveira@iffarroupilha.edu.br
RESUMO
Diante da importância da cultura da batata-doce, o presente estudo teve como objetivo otimizar a produção de plantas in vitro de diferentes cultivares. Segmentos nodais das cultivares ‘Abóbora’, ‘Catarina’, ‘Biaze’, ‘Da Costa’, ‘Americana’ e ‘Morada Inta’, foram cultivados em meio Murashige e Skoog e avaliados ao longo de oito semanas de cultivo in vitro. A cv. ‘Da Costa’ também foi cultivada em meio isento de fitorreguladores, em meio acrescido de benzilaminopurina (BAP) e/ou ácido giberélico (GA3) ou em sistema de cultivo fotoautotrófico. Constatou-se que a taxa de propagação é variável de acordo com o cultivar e com o período de tempo de cultivo. O intervalo entre os subcultivos foi dependente do índice de senescência das plantas. Na cv. ‘Da Costa’, uma maior produção de plantas completas foi obtida em meio nutritivo contendo BAP ou em meio isento de sacarose e em frascos vedados com algodão. As plantas oriundas de todas as cultivares apresentaram uma elevada taxa de sobrevivência durante a aclimatização.
Palavras-chave: batata-doce, Convolvulaceae, cv. ‘Da Costa’, cultivo fotoautotrófico, micropropagação.
ABSTRACT
Due to the importance of sweet potato culture, this study aimed to optimize the production of the in vitro plants of different cultivars. Nodal segments of cultivars ‘Abóbora’, ‘Catarina’, ‘Biaze’, ‘Da Costa’, ‘Americana’ e ‘Morada Inta’ were cultured on Murashige and Skoog medium and they were evaluated for eight weeks in an in vitro culture. The cv. ‘Da Costa’ was also grown on a medium free of plant growth regulators, supplemented with benzylaminopurine (BA) and/or gibberellic acid (GA3) or in photoautotrophic culture system. It was found that the propagation rate is variable, depending on the cultivar and according to time of cultivation. The interval among subcultures was dependent on the rate of plant senescence. In cv. ‘Da Costa’, a higher production of plants was obtained on medium containing BAP or on sucrose-free medium and in bottles sealed with cotton. Plants originating from all cultivars showed a high survival rate during the acclimatization.
Keywords: Convolvulaceae, cv. ‘Da Costa’, micropropagation, photoautotrofic culture, sweet potato.
Introdução
A batata-doce [Ipomoea batatas (L.) Lam.] é uma planta herbácea originária das Américas Central e do Sul. Devido as suas raízes tuberosas, é considerada uma hortaliça de grande importância econômica, sendo utilizada para comercialização in natura, para processamento em agroindústria e/ou produção de biocombustível (Silva et al., 2004).
Com relação ao valor nutricional, a batata-doce é uma excelente fonte de energia na forma de carboidratos (cerca de 30% da biomassa fresca), além de conter vários minerais e vitaminas. Em adição, os tubérculos apresentam, em média, 6% de proteína bruta por biomassa seca, além de elevados teores de carotenoides, os quais estão presentes nas cultivares de polpa amarela (Folquer, 1978; Martins et al., 1999).
O cultivo da batata-doce é muito difundido devido a sua facilidade de cultivo, rusticidade, colheita prolongada, baixo custo da produção, ampla adaptação às diferentes condições edafoclimáticas e multiplicidade de formas de aproveitamento (Murilo e Santos, 1999; Silva et al., 2008). Segundo dados da Food and Agriculture Organization (FAO, 2001), a batata-doce é cultivada em mais de 100 países, sendo que cerca de 90% da produção é obtida na Ásia, 5% na África e 5% no restante do mundo.
No Brasil, a batata-doce é considerada uma cultura de subsistência, sendo cultivada em todas as regiões, destacando-se os Estados do Nordeste, Sudeste e Sul. De acordo com o Atlas Socioeconômico do Rio Grande do Sul (RS) (2014), o RS é o maior produtor nacional, com uma produção média de 157.181 toneladas anuais, o que representa 31% da produção nacional. Apesar de, nos últimos anos, haver uma redução na área plantada tanto no Brasil como no RS, houve um aumento na produtividade da cultura, devido principalmente à evolução da tecnologia aplicada. Além disto, em plantas de propagação vegetativa, como a batata-doce, a qualidade fisiológica e sanitária das mudas é fundamental para a manutenção da produtividade da cultura (Arruda et al., 2003; Alam et al., 2010).
A batata-doce apresenta um grande número de cultivares, as quais se diferenciam principalmente pelo tamanho e cor das folhas, bem como pelo comprimento e espessura das ramas. Já os tubérculos podem variar em relação à posição na planta, cor da casca, cor da polpa e formato (Silva et al., 2004; 2008). Assim, em função das diferentes características dos tubérculos, cada cultivar é apropriada para determinadas aplicações. Por exemplo, na indústria alimentícia, há cultivares apropriadas para o consumo in natura, enquanto outras são mais adequadas para o preparo de pães, doces, compotas e/ou licores.
Tradicionalmente, a produção de mudas das diferentes cultivares de batata-doce é feita através de ramas (ou estacas) ou mediante a estimulação de gemas e brotos a partir das raízes tuberosas da planta (Folquer, 1978; Silva et al., 2008). Contudo, este tipo de propagação, apresenta uma série de desvantagens, como a alta proliferação de doenças, especialmente as viroses, micoplasmas, bactérias e fungos (Alam et al., 2010), as quais favorecem a depreciação dos tubérculos e reduzem significativamente a qualidade do produto e a produtividade da cultura.
Assim, tendo em vista a produção de mudas homogêneas e com alta qualidade fisiológica e sanitária, técnicas de cultura de tecidos (limpeza clonal através do isolamento de meristemas e micropropagação) vêm sendo muito utilizadas para a produção, em larga escala, de plantas matrizes de batata-doce (Wang e Hu, 1982; Castro e Andrade, 1995; Correa et al., 2003; Alam et al., 2010; Sivparsad e Gubba, 2012; Oliveira et al., 2013). A eficiência de um protocolo de micropropagação é influenciada por vários fatores, destacando-se o meio nutritivo, o qual deve ser ajustado para cada genótipo ou cultivar em estudo. Contudo, apesar de a batata-doce apresentar uma grande variabilidade genotípica, ainda há poucos estudos nesse sentido (Castro e Andrade, 1995; Oliveira et al., 2008). Em adição, são raras as informações sobre a produção in vitro da cultivar ‘Da Costa’, a qual apresenta expressiva importância econômica, devido a sua alta capacidade de acúmulo de biomassa e alta produtividade (Conceição et al., 2005).
Em geral, o cultivo da batata-doce pode ser conduzido em meio nutritivo isento de fitorreguladores (Oliveira et al., 2013), contudo dependendo da cultivar/genótipo, pode haver a necessidade de adição de fitorreguladores e/ou outros suplementos a fim de favorecer a produção de mudas (Castro e Andrade, 1995; Cassana et al., 2008, Oliveira et al., 2008; Alam et al., 2010; Sivparsad e Gubba, 2012). Assim, diante da importância da cultura, o presente estudo teve como objetivo comparar e otimizar a produção de mudas in vitro de seis cultivares de batata-doce, tendo em vista a produção de plantas de qualidade em escala comercial.
Material e Métodos
Como material vegetal utilizaram-se plantas assépticas e isentas de doenças, propagadas in vitro através do cultivo de meristemas. As plantas matrizes, pertencentes a seis cultivares (‘Abóbora’, ‘Catarina’, ‘Biaze’, ‘Da Costa’, ‘Americana’ e ‘Morada Inta’), foram cedidas pela Embrapa Clima Temperado, Pelotas, RS, Brasil.
As plantas foram multiplicadas, através de segmentos nodais (com cerca de 0,5 cm de comprimento) em meio MS (Murashige e Skoog, 1962), acrescido de 30 g L-1 de sacarose, 100 mg L-1 de mio-inositol e 7 g L-1 de ágar. O pH do meio foi ajustado para 5,8. As plantas foram mantidas em sala de crescimento sob temperatura de 25 ± 2°C, fotoperíodo de 16 horas de luz/8 horas de escuro e intensidade luminosa de cerca de 40 μmol m-2 s-1.
Plantas com quatro semanas de cultivo foram utilizadas como fornecedoras de explantes para os ensaios.
Potencial organogênico de seis cultivares de batata-doce in vitro
Segmentos nodais (com 0,5 cm de comprimento e com uma gema) das cultivares ‘Abóbora’, ‘Catarina’, ‘Biaze’, ‘Da Costa’, ‘Americana’ e ‘Morada Inta’, foram cultivados em meio MS, acrescido de 30 g L-1 de sacarose, 100 mg L-1 de mio-inositol e 7 g L-1 de ágar, sendo o pH ajustado para 5,8. O material foi mantido em sala de crescimento sob as mesmas condições de temperatura, fotoperíodo e intensidade luminosa descritas anteriormente.
As avaliações foram realizadas ao final de quatro semanas de cultivo mediante as seguintes variáveis: número de brotos e de segmentos nodais por planta, comprimento dos brotos (cm) e percentagem de enraizamento das plantas. Além disto, tendo em vista uma maior produção de mudas em relação ao tempo de permanência in vitro, as plantas foram avaliadas semanalmente, ao longo de oito semanas de cultivo, através da taxa de propagação (número de segmentos nodais produzidos por planta em um determinado intervalo de tempo).
Para a aclimatização, plantas completas, com quatro semanas de cultivo, foram transferidas para bandejas de isopor contendo o substrato comercial Plantamax Hortaliças® e mantidas em sala aclimatizada (25 ± 2°C, fotoperíodo de 16 horas e intensidade luminosa de cerca de 40 μmol m-2 s-1). A umidade do substrato foi mantida entre 45 e 60% da capacidade de campo, através de irrigações com água destilada. A taxa de sobrevivência foi avaliada após 30 dias do transplantio das plantas para o substrato. Após este período, as mudas foram transferidas para casa de vegetação.
Otimização da propagação clonal de batata-doce, cv. ‘Da Costa’
Tendo em vista as baixas taxas de propagação obtidas na cv. ‘Da Costa’, buscou-se otimizar a produção de mudas nesta cultivar através da instalação de dois ensaios.
a)Efeito do BAP e GA3 na propagação in vitro da cv. ‘Da Costa’
Segmentos nodais (com 0,5 cm de comprimento e com uma gema) foram cultivados em meio MS isento de fitorreguladores (tratamento controle) ou suplementado com BAP (1,0 mg L-1) em conjunto ou não com GA3 (0,1 ou 0,5 mg L-1). Após quatro semanas, as plantas foram avaliadas mediante o número de segmentos nodais e comprimento dos brotos (cm). Após, o material foi subcultivado em meio MS isento de fitorreguladores para enraizamento. O percentual de enraizamento das plantas e o número de raízes por planta foram avaliados após quatro semanas de cultivo. As plantas foram mantidas em sala de crescimento sob as mesmas condições de temperatura, fotoperíodo e intensidade luminosa descritas anteriormente.
Para aclimatização, plantas completas, com cinco semanas de cultivo, foram transferidas para bandejas de isopor contendo o substrato comercial Plantamax Hortaliças® e mantidas em sala aclimatizada (25 ± 2°C, fotoperíodo de 16 horas e intensidade luminosa de cerca de 40 μmol m-2s-1). A umidade do substrato foi mantida entre 45 e 60% da capacidade de campo, através de irrigações com água destilada. A taxa de sobrevivência foi avaliada após quatro semanas do transplantio das plantas para o substrato.
b) Efeito da sacarose e vedamento do frasco na propagação in vitro da cv. ‘Da Costa’
Segmentos nodais (com cerca de 0,5 cm de comprimento e com uma gema) foram cultivados em meio MS acrescido de 100 mg L-1 de mio-inositol e 7 g L-1 de ágar. Testaram-se duas concentrações (0 e 30 g L-1) de sacarose e três tipos de vedamento (algodão, plástico parafilme ou papel alumínio) do frasco de cultivo, totalizando seis tratamentos. O pH do meio foi ajustado para 5,8.
As plantas foram mantidas em sala de crescimento, sendo avaliadas após quatro semanas da instalação do ensaio, mediante a percentagem de regeneração de brotos e de raízes.
Análise dos dados
O delineamento experimental foi o inteiramente ao acaso com seis repetições, sendo cada repetição formada por cinco plantas. Os dados foram submetidos à análise de variância e analisados pelo teste de Tukey em nível de 1% de probabilidade de erro.
Resultados e discussão
Potencial organogênico de seis cultivares de batata-doce in vitro
Nos dias atuais, devido à expressiva importância da batata-doce, vários estudos vêm sendo conduzidos com diferentes cultivares, tendo em vista a otimização de propagação clonal de mudas, bem como a conservação de genótipos in vitro (Oliveira et al., 2008; Sivparsad e Gubba, 2012; Arrigoni-Blank et al., 2014). No presente estudo, todas as cultivares apresentaram bom potencial organogênico, formando plantas completas após quatro semanas de cultivo (Quadro 1). De fato, estudos recentes conduzidos por Alam et al. (2010), ressaltaram que em batata-doce, muitas vezes, a multiplicação in vitro de brotos ocorre concomitantemente ao enraizamento, não sendo necessária a adição de suplementos ao meio nutritivo para induzir a regeneração de raízes.
Não foi observada diferença estatística em relação ao número de brotos entre as cultivares estudadas, contudo estas diferiram em relação ao número de segmentos nodais, comprimento e regeneração de raízes (Quadro 1), o que concorda com Sivparsad e Gubba (2012), de que o potencial para formar mudas in vitro em batata-doce é genótipo-específico.
Em batata-doce, brotos apicais ou nodais são os mais indicados para a produção de mudas in vitro (Sivparsad e Gubba, 2012). Entretanto, a baixa formação de brotos obtida neste estudo se justifica pela presença de apenas uma gema por segmento nodal e, especialmente, pelo uso de um meio nutritivo isento de fitorreguladores. Neste caso, a indução de brotos ocorre pela quebra da dominância apical, que estimula o desenvolvimento das gemas em decorrência do balanço hormonal endógeno do explante ser favorável às citocininas. Em plantas propagadas in vitro dessa forma, a variável que reflete diretamente o potencial de cada genótipo em formar novas mudas em um determinado intervalo de tempo é a taxa de propagação (expressa pelo número de segmentos nodais ou microestacas). Em adição, segundo Alam et al. (2010), o desenvolvimento de plantas de batata-doce a partir de gemas pré-formadas, evitando-se a regeneração a partir de calos ou via gemas adventícias, é importante para evitar distúrbios fisiológicos e variação somaclonal.
Além do fato de a taxa de propagação de mudas ser variável de acordo com a cultivar estudada (Quadro 1), também constatou-se que esse parâmetro varia conforme o tempo de cultivo (Figura 1A). Estes resultados complementam os estudos de Castro e Andrade (1995), Oliveira et al. (2008) e Sivparsad e Gubba (2012).
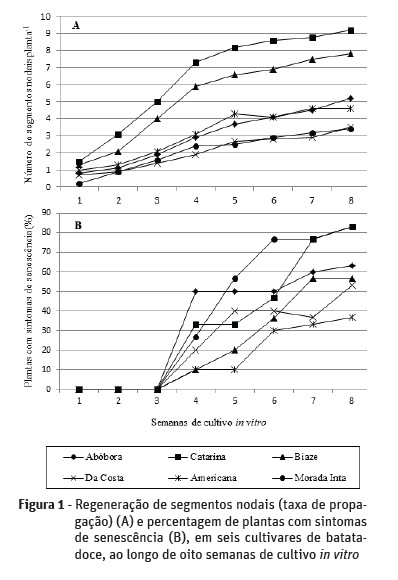
Apesar de todas as cultivares apresentarem um aumento no número de segmentos nodais ao longo do tempo, constatou-se que na quarta semana de cultivo in vitro, as plantas começaram a apresentar sintomas de senescência nas folhas, cuja frequência variou conforme a cultivar (Figura 1B). Estes sintomas caracterizaram-se por clorose, amarelecimento e/ou abscisão de folhas. Estes dados mostram-se importantes tendo em vista a determinação de um maior intervalo possível entre os subcultivos que culmine na produção de um maior número de mudas sem, contudo, prejudicar a qualidade fisiológica das plantas.
As cultivares ‘Catarina’ e ‘Biaze’ destacaram-se por apresentarem maior taxa de propagação in vitro (Figura 1A) em relação às demais cultivares. Ambas as cultivares também apresentaram plantas com maior comprimento e percentual de enraizamento (Quadro 1). O percentual de plantas com sintomas de senescência nessas cultivares ficaram mais evidentes (mais de 50% das plantas) a partir da sexta semana de cultivo in vitro, indicando que tanto a cv. ‘Catarina’ como a cv. ‘Biaze’ podem ser subcultivadas a cada seis semanas, com taxas de propagação de 8,6 e 6,9, respectivamente (Figura 1A e 1B).
Nas cultivares ‘Abóbora’ e ‘Da costa’, a taxa de propagação das plantas aumentou significativamente (P ≤ 0,05) ao longo do período de cultivo in vitro. Entretanto, a cv. ‘Abóbora’ apresentou sintomas de senescência após quatro semanas de cultivo, cujo valor permaneceu estável até a sexta semana. Assim, sugere-se que essa cultivar seja subcultivada a cada seis semanas, com uma taxa de propagação de 4,1 (Figura 1A e 1B).
Nas cultivares ‘Catarina’, ‘Biaze’, ‘Americana’ e ‘Morada Inta’, não foi observado um incremento significativo (P ≥ 0,05) na taxa de propagação de mudas a partir da quinta semana de cultivo (Figura 1A). Em relação à senescência, constatou-se que as cultivares ‘Da Costa’ e ‘Americana’ apresentaram menores taxas de senescência em relação às demais cultivares, podendo ser subcultivadas em intervalos de sete e oito semanas de cultivo (Figura 1B), com taxas de propagação de 2,9 e 4,6, respectivamente.
Por outro lado, na cultivar ‘Morada Inta’, a percentagem de plantas com sintomas de senescência cresceu rapidamente, sendo sugerido que esta cultivar seja subcultivada a cada quatro semanas de cultivo (Figura 1B). Esta cultivar, juntamente com a ‘Da Costa’, também apresentou as menores taxas de propagação (Quadro 1, Figura 1A).
Dentre os fatores que favorecem a senescência das plantas in vitro, destacam-se a redução dos nutrientes e de água no meio nutritivo e o acúmulo de gases, especialmente o etileno. Na maioria das espécies, o ideal é que um novo subcultivo seja realizado antes do início da fase de senescência. No presente estudo, verificou-se que as cultivares responderam de forma diferente em relação ao início e progressão da senescência, indicando que esta variável é genótipo-específica. Assim, os intervalos ideais entre cada subcultivo devem variar conforme a cultivar, reduzindo-se, assim, os custos com reagentes, energia, vidraria, além da mão de obra e do manuseio da cultura.
No presente estudo, todas as cultivares produziram plantas completas com brotações alongadas, sistema radicular bem desenvolvido e morfologia normal. Independente do tempo de cultivo, as maiores taxas de propagação in vitro foram registradas nas cultivares ‘Catarina’ e ‘Biaze’, as quais apresentaram maior vigor, maior número de segmentos nodais, plantas mais alongadas e melhores taxas de enraizamento em relação às demais cultivares. Assim, sugere-se o uso do meio nutritivo MS, isento de fitorreguladores, para a propagação clonal in vitro dessas cultivares, as quais podem ser subcultivadas a cada seis semanas (Figura 1A). Por outro lado, as cultivares ‘Abóbora’, ‘Da Costa’, ‘Americana’ e ‘Morada Inta’ apresentaram uma menor produção de segmentos nodais (Quadro 2), indicando a necessidade de estudos subsequentes no sentido de favorecer a produção de mudas in vitro, bem como de estratégias para minimizar os processos de senescência (Figura 1A e 1B). Segundo Castro e Andrade (1995) e Oliveira et al., (2008), muitas vezes, faz-se necessário testar diferentes composições de meio nutritivo no sentido de otimizar a produção de mudas nas diferentes cultivares.
Durante o processo de aclimatização, as plantas produzidas in vitro apresentaram uma elevada taxa de sobrevivência. Exceto na cv. ‘Americana’, onde 83,3% das mudas sobreviveram, nas demais cultivares, observou-se 100% de sobrevivência após o plantio. Todas as plantas produzidas in vitro apresentaram crescimento e desenvolvimento normais. Outros estudos conduzidos com batata-doce também obtiveram sucesso durante a fase de aclimatização das mudas (Sivparsad e Gubba, 2012).
Otimização da propagação clonal de batata-doce, cv. ‘Da Costa’
Efeito da BAP e GA3 na propagação in vitro
Os explantes apresentaram uma alta taxa de regeneração independente do meio nutritivo. Os melhores resultados em relação à taxa de propagação (expressa pelo número de segmentos nodais) foram obtidos com o uso da BAP. Por outro lado, a adição do GA3 ao meio não favoreceu de forma significativa o desenvolvimento das plantas (Quadro 2).
Os resultados obtidos com a BAP foram significativamente superiores ao controle (meio isento de fitorreguladores) (Quadro 1), uma vez que as plantas apresentaram um aumento considerável no número de segmentos nodais, crescimento dos brotos e, especialmente, em relação ao enraizamento (Quadro 2). Assim, o uso da BAP poderá ser uma alternativa viável para o incremento da taxa de multiplicação da cv. ‘Da Costa’, viabilizando a produção de plantas completas nesta cultivar. A BAP também foi benéfica para a multiplicação in vitro de batata-doce, cv. ‘Blesbok’ (Sivparsad e Gubba, 2012).
Citocininas, como a BAP, são muito utilizadas durante a fase de multiplicação in vitro de batata-doce, pois favorecem a propagação de gemas e brotos (Sivparsad e Gubba, 2012), o que corrobora os resultados obtidos no presente estudo com a cv. ‘Da Costa’, onde o uso da BAP induziu um maior número de segmentos nodais. Além dos efeitos benéficos na regeneração de brotos, foi observado que, em algumas espécies, as citocininas também inibem a senescência e a abscisão foliar in vitro (Oliveira et al., 2007), efeitos fisiológicos comuns desses hormônios nas plantas in vivo. Na cv. ‘Da Costa’, os primeiros sintomas de senescência já foram visíveis (em 20% das plantas) na quarta semana de cultivo em meio MS isento de fitorreguladores (Figura 1B). Já na presença da BAP, esses sintomas não foram visíveis (dados não mostrados).
No entanto, segundo George (1996), o uso de citocininas em concentrações não adequadas para o genótipo em estudo, pode induzir distúrbios fisiológicos, como redução no alongamento dos brotos e inibição do enraizamento. Na cv. ‘Da Costa’, ao contrário, o uso da BAP favoreceu o crescimento das plantas (Quadro 2).
Em batata-doce, uma das estratégias adotadas para minimizar os efeitos da BAP no enraizamento das mudas é a transferência dos brotos para um meio nutritivo isento de regulador de crescimento (Sivparsad e Gubba, 2012). De fato, neste estudo com a cv. ‘Da Costa’, a transferência dos brotos induzidos com BAP para um meio isento de fitorreguladores foi essencial para o desenvolvimento do sistema radicular (Quadro 2).
Os efeitos fisiológicos do GA3 nas plantas são bem conhecidos, sendo que na cultura de tecidos o ácido giberélico vem sendo utilizado para induzir alongamento dos brotos e, desta forma, favorecer a propagação in vitro (Isogai et al., 2008). Alam et al., (2010) observou que na propagação clonal de batata-doce, o meio contendo citocinina e GA3 foi o mais efetivo. Contudo, ao contrário, o GA3 não favoreceu significativamente a produção de mudas da cv. ‘Da Costa’ (Quadro 2).
As mudas foram aclimatizadas e, após 30 dias do transplantio, registrou-se um índice de 80% de sobrevivência. Desta forma, o cultivo de segmentos nodais de batata-doce, cv. ‘Da Costa’, em meio acrescido de 1,0 mg L-1 de BAP, seguido do subcultivo dos brotos em meio isento de fitorreguladores mostrou-se uma metodologia viável para a micropropagação dessa cultivar, devido a alta taxa de multiplicação, bom desenvolvimento da parte aérea e raízes e uma boa adaptação às condições ex vitro.
Efeito da sacarose e vedamento do frasco na propagação in vitro
Em relação ao percentual de regeneração de brotos não foi verificado interação entre concentração de sacarose e o tipo de vedamento do frasco, contudo ambos os fatores foram estatisticamente significativos. Os maiores percentuais de regeneração de brotos foram registrados em meio suplementado com sacarose e em frascos de cultivo vedados com algodão (Quadro 3). Conforme salientam Marino et al. (2010), a sacarose afeta significativamente as respostas fisiológicas das plantas, atuando tanto como fonte de energia/esqueletos de carbono, bem como na regulação osmótica do meio de cultura.
Quanto ao percentual de plantas enraizadas, constou-se uma interação significativa entre sacarose e o tipo de vedamento do frasco de cultivo, sendo os melhores resultados registrados nas plantas cujos frascos foram vedados com algodão, não havendo diferença estatística entre as concentrações de sacarose (Quadro 4).
Em batata-doce, bem como na maioria das culturas, a micropropagação é feita em condições heterotróficas ou fotomixotróficas, onde os explantes são cultivados em frascos hermeticamente ou semi-hermeticamente fechados, impedindo que ocorram trocas gasosas entre o ambiente interno e externo. Assim, esta forma de vedamento dos frascos de cultivo não favorece a atividade fotossintética das plantas in vitro. Neste caso, faz-se necessário adicionar ao meio de cultivo uma fonte de carbono, sendo a sacarose o açúcar mais utilizado na micropropagação de batata-doce (Alam et al., 2010). Isto explica, por que neste estudo, quando se utilizou tipos de vedação semi-hermética (plástico parafilme e papel alumínio), que não favorecem as trocas gasosas, as melhores respostas em relação à organogênese de raízes ocorreu em meio suplementado com sacarose (Quadro 4).
Atualmente, uma das estratégias mais promissoras para melhorar as respostas in vitro é a micropropagação em meio isento de sacarose e sob condições que promovam a indução da capacidade fotossintética (cultivo fotoautotrófico), visando a produção endógena dos carboidratos necessários para o crescimento e desenvolvimento (Afreen et al., 2002). Uma das formas de estimular a condição autotrófica in vitro é o uso de um sistema de vedação que promova as trocas gasosas, como o algodão utilizado no presente estudo.
Outros trabalhos vêm ressaltando a importância de as mudas micropropagadas de batata-doce apresentarem um bom desempenho fotossintético quando cultivadas em campo (Arruda et al., 2003; Cassana et al., 2008), sendo que este desempenho depende muito das condições de cultivo in vitro.
No presente estudo, com a cv. ‘Da Costa’, os resultados obtidos em relação à regeneração de brotos e raízes deve-se, possivelmente, ao uso do algodão como forma de vedação do frasco de cultivo. Este tipo de vedação, possivelmente, favoreceu as trocas gasosas, levando a uma intensificação da atividade fotossintética que culmina na síntese de carboidratos necessários à organogênese (formação de brotos e raízes) pela própria planta.
Esta metodologia de cultivo é pioneira em batata-doce e apresenta inúmeras vantagens, como redução de custos e uma melhor qualidade morfofisiológica das plantas regeneradas. A batata-doce, em especial a cv. ‘Da Costa’, é uma cultura muito expressiva pois apresenta um custo baixo de produção, alto potencial produtivo e grande valor alimentício. Os resultados obtidos no presente estudo otimizaram e viabilizam com sucesso a produção de mudas de qualidade e em larga escala da cv. ‘Da Costa’.
Conclusões
A taxa de propagação de batata-doce varia de acordo com a cultivar e com o período de tempo de cultivo, sendo que as cultivares ‘Catarina’ e ‘Biaze’ apresentam maior potencial para a produção de mudas in vitro em relação as cultivares ‘Abóbora’, ‘Da Costa’, ‘Americana’ e ‘Morada Inta’.
O intervalo entre os subcultivos depende do índice de senescência das plantas, variando de quatro-oito semanas de acordo com a cultivar.
Uma maior produção de mudas completas da cv. ‘Da Costa’ é obtida em meio nutritivo contendo 1,0 mg L-1 de BAP ou, alternativamente, as mudas podem ser produzidas em sistema de cultivo fotoautotrófico.
As mudas apresentam uma elevada taxa de sobrevivência durante a aclimatização.
Agradecimentos
Os autores agradecem à Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS, RS, Brasil) e ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brasil), pelo auxílio financeiro.
Referências Bibliográficas
Afreen, F.; Zobayed, S.M.A. e Kozai, T. (2002) - Photoautotrophic culture of Coffean arabusta somatic embryos: photosynthetic ability and growth of different stage embryos. Annals of Botany, vol. 90, n. 1, p. 11-19. [ Links ]
Alam, I. et al. (2010) - Effect of growth regulators on meristem culture and plantlets establishment in sweet potato [Ipomoea batatas (L.) Lam.]. Plant Omics Journal, vol. 3, n. 2, p. 35-39. [ Links ]
Arigoni-Blank, M.F. et al. (2014) - In vitro conservation of sweet potato genotypes. The Scientific World Journal, vol. 2014, p. 1-7. [ Links ]
Arruda, A. da S. et al. (2003) - Efeitos do benomyl e da kasugamicina na produção da batata doce micropropagada. Bioscience Journal Uberlândia, vol. 19. n. 2, p. 99-103. [ Links ]
Atlas socioeconômico do Rio Grande do Sul. (2014) - [citado 2014-09-20]. Disponível em: http://www.scp.rs.gov.br/atlas/conteudo.asp?cod_menu_filho=819&cod_menu=817&tipo_menu=ECONOMIA&cod_conteudo=1502 [ Links ]
Cassana, F.F. et al. (2008) - Atividade fotoquímica máxima do fazer fotossistema II em plantas de batata-doce cultivadas in vitro e aclimatizadas. Revista Brasileira de Agrociência, vol. 14, n. 4, p. 46-51. [ Links ]
Castro, O.F.A. e Andrade, A.G. (1995) - Cultura de meristemas de batata-doce [Ipomoea batatas (L.) Lam.]. Pesquisa Agropecuária Brasileira, vol. 30, n. 7, p. 917-922.
Conceição, M.K. et al. (2005) - Análise de crescimento de plantas de batata-doce [Ipomoea batatas (L.) Lam.] cultivares abóbora e da costa. Revista Brasileira de Agrociência, vol. 11, n. 3, p. 273-278. [ Links ]
Corrêa, R.M. et al. (2003) - Potencial do carvão ativado, filtro amarelo e interação fotoperíodo/temperatura na formação de raízes tuberosas de batata-doce in vitro. Ciência Rural, vol. 33, n. 3, p. 423-430. [ Links ]
FAO (Food and Agriculture Organization of the United Nations). 2001 – [citado 2014-05-09]. Disponível em: http://apps.fao.org
Folquer, F. (1978) - La batata (camote): estudio de la planta y su produccion comercial. San José, Hemisfério Sul, 134 p. [ Links ]
George, E. (1996) - Plant propagation by tissue culture: the tecnology. Eversley, Exegetics Limited, 1996. 758 p. [ Links ]
Isogai, S.; Touno, K. e Shimomura, K. (2008) - Gibberellic acid improved shoot multiplication in Cephaelis ipecacuanha. In vitro Cellular & Development Biology-Plant,vol. 44, p. 216-220. [ Links ]
Marino, G. et al. (2010) - Effect of carbohydrates on in vitro low-temperature storage of shoot cultures of apricot. Scientia Horticulturae, vol. 126, p. 434-440. [ Links ]
Martins, M.; Pinto, J.E.B.P. e Paiva, E. (1999) - Purificação e produção de anticorpos de esporamina: uma proteína específica da batata-doce. Ciência e Agrotecnologia, vol. 23, n. 4, p. 849-855. [ Links ]
Murashige, T. e Skoog, F. (1962) - A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum, v. 15, p. 473-497. [ Links ]
Murilo, D.V. e Santos, J.E. (1999) - Avaliação de clones de batata-doce quanto à resistência a insetos do solo. Revista Caatinga, vol. 12, n. 1-2, p. 13-16. [ Links ]
Oliveira, C.M. et al. (2013) - Efeito de reguladores de crescimento na micropropagação in vitro de batata doce. Global Science and Technololgy, vol. 06, n. 03, p. 108-115. [ Links ]
Oliveira, L.M.; Paiva, R; Santana, J.R.F.; Nogueira, R.C.; Soares, F.P. e Silva, L.C. (2007) – Efeito de citocininas na senescência e abscisão foliar durante o cultivo in vitro de Annona glabra L. Revista Brasileira de Fruticultura, vol. 29, n. 1, p. 25-30.
Oliveira, M.K.T.; Neto, F.B.; Câmara, F.A.; Dombroski, J.L.D. e Freitas, R.M.O. (2008) -Multiplicação in vitro de batata-doce (Ipomoea batatas Lam.). Revista Caatinga, vol. 21, n. 4, p. 129-134. [ Links ]
Silva, J.B.C.; Lopes, C.A. e Magalhães, J.S. (2004) - Cultura da batata-doce (Ipomoea batatas Lam.), n. 6 [citado 2014-10-11). Disponível em: http://www.cnph.embrapa.br/sistprod/batatadoce [ Links ]
Silva, J.B.C.; Lopes, C.A. e Magalhães, J.S. (2008) - Batata-doce (Ipomoea batatas). [citado 2014-10-11] Disponível em: http://www.cnph.embrapa.br/paginas/sistemas_producao/cultivo_batata_doce.htm [ Links ]
Sivparsad, B.J. e Gubba, A. (2012) – Development of an efficient plant regeneration protocol for sweet potato (Ipomoea batatas L.) cv. Blesbok. African Journal of Biotechnology, vol. 11, n. 84, p. 14982-14987.
Wang, P. e Hu, C. (1982) - In vitro mass tuberization and virus-free seed-potato production in Taiwan. American Potato Journal, vol. 59, n. 1, p. 33-37. [ Links ]
Recebido/Received: 2014.12.03
Aceite/accepted: 2015.02.16

