










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.38 no.3 Lisboa set. 2015
ARTIGO
Incidência de Bipolaris sorokiniana nas sementes e transmissão para plantas de cevada
Incidence of Bipolaris sorokinianain seeds and transmission to barley plants
Josiane T. Ferrari1* e Edilberto Possamai2
1,* Instituto Biológico, CPDSV/Laboratório de Doenças Fúngicas em Horticultura, Av. Conselheiro Rodrigues Alves, 1252, Vila Mariana, CEP 04014-002, São Paulo, SP Brasil. E-mail: takassaki@biologico.sp.gov.br, author for correspondence
2 Departamento de Fitotecnia e Fitossanitarismo, UFPR/SCA, R. dos Funcionários, 1540, Juvevê, CEP 80035-050, Curitiba, PR. Brasil. E-mail: possamai@ufpr.br
RESUMO
O potencial de rendimento da cevada é diminuído principalmente pela ocorrência de doenças como a mancha marrom (Bipolaris sorokiniana) transmitida pelas sementes. O objetivo desse trabalho consistiu em determinar a correlação entre diferentes níveis de incidência do fungo em sementes e sua transmissão para plantas de cevada em campo e estufa e avaliar a capacidade de transmissão do fungo pelas sementes. Sementes de vários lotes da cultivar ‘BR 2’ com níveis de incidência de 2,7; 7,5; 12,5; 16,5; 31,0 e 55,5%, previamente selecionados em laboratório, foram semeadas em campo, em quatro repetições. As avaliações foram realizadas aos sete dias após a emergência, contando-se o número de plantas com sintomas de helmintosporiose nas folhas e determinando-se a porcentagem em relação à emergência. Em estufa, as sementes foram semeadas em caixas plásticas com aproximadamente 13 kg de solo esterilizado. Foram colocadas 400 sementes a 3 cm de profundidade. Constatou-se a importância da semente infectada como fonte de inóculo primário. Observou-se que a transmissão do patógeno no campo foi mais eficiente quando o nível de incidência foi de 55,5%. Em estufa, houve correlação apenas entre a incidência nas sementes e nas raízes. Concluiu-se que não houve correlação entre diferentes níveis B. sorokiniana nas sementes e a sua incidência em campo e que níveis de 55,5% diminuíram a emergência das plantas no campo e em estufa.
Palavras-chave: fungo, Hordeum vulgare, incidência, taxa de transmissão, ponta preta
ABSTRACT
The yeld potential of barley is reduced mainly because of diseases such as spot blotch caused by Bipoloaris sorokiniana which is seed transmitted. The objectives of the work were to study the correlation among different levels of seed infection and its transmission to barley plants under field and greenhouse conditions, and determine the capacity of seed-transmission of the fungus. Barley seeds cv ‘BR 2’ naturally infected with the levels of 2.7; 7.5; 12.5; 16.5; 31.0 and 55.5% of the fungus were sown in the field, with four replications. The evaluation of disease seedling showing simptoms of helminthosporiosis and its percentage in relation to the seed emergence, were made seven days after emergence. In greenhouse, seeds were sown in plastic trays containing with aproximatelly 13 kg of soil previously sterelized. Four hundred seeds of each tretament (infection level) were planted and covered with 3 cm of soil (3 cm depth). The importance of the infected seeds as a source of primary inoculum was demonstred. It was observed that on the field the transmission of the pathogen was more efficient when the seed infection level was 55%. On the other hand, in greenhouse there was correlation only between seed infection and roots. In conclusion, there was no correlation among the seed infection levels of B. sorokiniana and their incidence in the field, and seed infection levels of 55% reduce seedling emergence both, in the field or in greenhouse.
Key words: black point, fungus, Hordeum vulgare, infection, seed, transmission.
Introdução
A produção mundial de cevada, em 2005, foi de 140 milhões de toneladas (Barley (Hordeum) Maps and Statistics, 2007), da qual cerca de 13%, ou seja, aproximadamente 18 milhões de toneladas de cevada (Hordeum vulgare) foram utilizadas para a produção de malte cervejeiro. A cultura, no Brasil, atualmente, é feita na região Sul e nos estados de Goiás, de Minas Gerais e de São Paulo (De Mori e Minella, 2012). Em 2011, a área semeada de cevada no Brasil foi de 88,4 mil hectares com uma produção de 305,1 mil toneladas (Conab, 2014).
A cevada, no Brasil, está sujeita a uma série de doenças causadas por fungos e, pelo menos oito desses patógenos podem ser transmitidos por sementes. Todas as doenças fúngicas, além de causarem perdas de rendimento, afetam a qualidade comercial da cevada, destacando-se entre elas a mancha marrom, conhecida também como helmintosporiose, que provoca lesões nas folhas, de formato oval ou alongadas de 1 a 2 mm, com coloração marrom variando de pardo a escuro circundadas por um halo amarelo e margem definida variando em tamanho. As manchas podem aumentar e coalescer cobrindo grandes áreas das folhas, disseminando-se entre os nós e entrenós. Quando ataca as espigas, causa descoloração e ponta preta nos grãos que irá influenciar a qualidade do malte e cerveja (Arias, 1995). A doença é causada pelo fungo Cochliobolus sativus (Ito e Kurib.), Drechs. ex. Dastur, anamorfo: Bipolaris sorokiniana (Sacc. in. Sorok) Shoem. Esse patógeno apresenta a característica de utilizar como substrato todos os órgãos dos cereais de inverno. Dessa forma decorrem duas fases distintas da doença: interferência no processo fotossintético, quando o ataque ocorre nos órgãos verdes como folhas, bainhas, colmos, glumas, aristas e sementes em formação e interferência na busca e absorção de nutrientes e água, que constitui a fase da doença que ocorre em órgãos subterrâneos como raízes seminais, mesocótilos, raízes secundárias e coroa (Reis, 1981).
Um outro tipo de doença causada por B. sorokiniana em cevada é a podridão comum de raízes, caracterizado por lesões pequenas, ovais e marrons sobre as raízes primárias e secundárias, no mesocótilo (também chamado de entre-nó subcoronal) e na coroa das plantas e são originadas de conídios presentes no solo ou do micélio presente na semente. Se o inóculo estiver presente nas sementes, as plântulas podem morrer ou ficarem subdesenvolvidas, com escurecimento das raízes e do coleóptilo. As lesões podem invadir todo o sistema radicular estendendo-se à base do colmo (Picinini e Fernandes, 1995). Quando severamente atacadas, as raízes são pouco desenvolvidas e as plantas afilham pouco.
O controle mais eficaz para essa doença é a utilização de sementes sadias, pois o fungo é transmitido por sementes e o tratamento com fungicidas só é viável com níveis de infecção abaixo de 40%. A da rotação de culturas é outra prática de controlo, visando a redução do inoculo na palha.
O fungo B. sorokiniana está associado à semente de cevada de duas formas, aderido externamente, e nesse caso, diz-se que a semente está infestada sendo a taxa de transmissão muito baixa. Outra forma de associação é quando o fungo está localizado internamente na semente na forma de micélio no pericarpo e no endosperma, nesse caso a taxa de transmissão é mais elevada, garantindo eficientemente a sobrevivência do patógeno e sua posterior passagem aos órgãos radiculares e aéreos (Reis e Casa, 1998).
Lotes de sementes de cevada apresentando nível de infecção com B. sorokiniana acima de 5% são suficientes para produzirem o inóculo necessário ao desenvolvimento de uma epidemia no campo, caso as condições de ambiente sejam favoráveis (Picinini e Fernandes, 1995).
É importante conhecer o modo de transmissão dos fitopatógenos pela semente, para determinar como esta atuará como fonte de inóculo e como a disseminação poderá ocorrer. Entretanto, para a maioria dos patógenos de sementes as pesquisas sobre danos por eles ocasionados são poucas ou inexistentes (Lasca, 1997).
O objetivo desse trabalho consistiu em avaliara a correlação entre diferentes níveis de incidência de Bipolaris sorokiniana em sementes e sua taxa de transmissão para plantas de cevada em campo e em estufa.
Material e Métodos
Teste de germinação
Foram selecionadas amostras de sementes aparentemente doentes, isto é, com sintomas de ponta preta, para realização dos testes de germinação e determinação do nível de incidência de B. sorokiniana. Para esses teste foram selecionadas oito amostras de sementes de cevada da cultivar ‘BR-2’, suscetível à helmintosporiose, não tratadas com fungicidas. As amostras foram provenientes de campos de produção de sementes dos municípios de Passo Fundo, RS, Papanduva, SC e Lapa, PR.
O teste de germinação foi realizado de acordo com as normas recomendadas (Brasil, 2009), utilizando rolo de papel como substrato. Para cada amostra foram utilizadas 400 sementes, divididas em 100 sementes por repetição. Essas sementes foram distribuídas em 8 rolos de papel com 50 sementes cada. Para cada rolo, foram utilizadas 3 folhas de papel germitest previamente umedecidas em água destilada. As sementes foram colocadas em câmara de germinação à temperatura de 20 ºC e às escuras. A germinação foi avaliada após 7 dias, considerando as plântulas normais, anormais e mortas.
Teste para determinação da incidência de Bipolaris sorokoniana em semente Paralelamente ao teste de germinação, para determinação do nível de incidência dessas amostras, utilizou-se o método de papel de filtro (‘blotter test’), conforme Neergard (1979). De cada lote foram analisadas 400 sementes por amostra, totalizando oito tratamentos, com quatro repetições de 100 sementes e observada a incidência (%) de B.sorokiniana. O delineamento estatístico utilizado foi o de blocos inteiramente casualizados. Para esse método, foram utilizadas caixas plásticas (gerbox), tratadas previamente com hipoclorito de sódio 1%. Em cada placa foram colocadas 20 sementes, totalizando cinco placas por repetição e 20 placas por tratamento. As placas foram mantidas em BOD à temperatura de 20 ºC sob fotoperíodo de 12-h. Após sete dias procedeu-se à avaliação da incidência de B. sorokiniana, nas sementes, observando-se as estruturas do fungo, ou seja, conidióforos e conídios com auxílio de microscópios estereoscópico e óptico.
Dos oito tratamentos iniciais, foram selecionados seis diferentes níveis de incidência do fungo nas sementes para o ensaio em campo e em estufa.
Ensaios de campo
Os ensaios foram instalados dentro do período preferencial de sementeira da cevada cervejeira no município da Lapa, PR , localizado a 25º46'02" Latitude Sul e 49º43'10" Longitude Oeste e a 907 m de altitude. A área escolhida ficou sem a cultura ou com a cultura de cevada por aproximadamente 18 meses, período esse recomendado para o controle de doenças do sistema radicular da cevada, pela redução do potencial de inóculo nos restos culturais (Santos et al., 1995). As operações culturais obedeceram às recomendados para a cultura, bem como a observação de outras doenças e pragas durante o período de realização do estudo.
Na determinação da taxa de transmissão de b. sorokiniana da semente para a planta epara cada nível de incidência determinado em laboratório, foram testadas 2000 sementes no campo. As parcelas foram constituídas de seis fileiras espaçadas de 0,17 m entre si e com 3,00 m de comprimento, com área total igual a 2,55 m2 contendo cada fileira 83 sementes, totalizando 498 sementes por parcela. A cultura foi instalada no terreno por recurso a sementeira direta, em área cultivada anteriormente com milho.
A emergência ocorreu oito dias após a sementeira e a leitura final foi feita 13 dias após a sementeira, contando-se as plantas das quatro fileiras centrais de cada parcela.
Para a determinação do percentagem de plantas com B. sorokiniana, as plantas das 4 fileiras centrais das parcelas foram avaliadas pela contagem de plantas doentes, observando-se a presença de sintomas na plúmula ou primeira folha aos sete dias após a emergência e posteriormente a cada quinze dias até a época de colheita.
O delineamento experimental utilizado foi o de blocos casualizados com seis tratamentos (níveis de incidência de 2,7, 7,5, 12,5, 16,5, 31,0 e 55,5%) e quatro repetições por tratamento. Após a colheita e secagem, amostras de sementes de cada parcela do ensaio foram pesadas, para determinação do rendimento.
Essas amostras foram submetidas a testes de sanidade pelo método do papel de filtro (‘blotter test’), para determinar a porcentagem de sementes infectadas por B. sorokiniana. No período de realização do experimento foram registados os dados de temperatura máxima e mínima, bem como a precipitação para observar a influência das condições climáticas sobre o desenvolvimento do patógeno nas plantas.
Transmissão de B. sorokiniana da planta para a semente no campo
Após a colheita no campo, limpeza, secagem e pesagem, as amostras de sementes de cada parcela do experimento, foram submetidas a testes de germinação em rolo de papel germitest e de sanidade, pelo método do papel de filtro (“blotter test”), para determinar a existência de sementes infectadas por B. sorokiniana. As amostras foram semeadas conforme descrito anteriormente. O delineamento utilizado foi o de blocos inteiramente casualizados com seis tratamentos e quatro repetições, com 100 sementes por repetição.
A contagem de plântulas foi feita aos 7 e 13 dias após a emergência. Após 40 dias as plantas foram cuidadosamente retiradas do substrato, lavadas e avaliadas quanto a presença de sintomas nos coleóptilo e raízes.
Ensaios em estufa
Transmissãode B. sorokiniana da semente para a plântula: para a determinação da taxa de transmissãode B. sorokiniana da semente para a plântula foram realizados dois ensaios: o primeiro foi feito a partir das amostras selecionadas em laboratório e que serviram para os ensaios de campo e o segundo a partir das sementes colhidas provenientes dos ensaios de campo. Os experimentos foram conduzidos sob temperatura aproximada de 25 ºC.
Transmissãode B. sorokiniana da semente para a plântula: para a determinação da taxa de transmissão de b. sorokiniana da semente para a planta,seis amostras com diferentes níveis de incidência de B.sorokinana: 2,7, 7,5, 12,5, 16,5, 31,0 e 55,5%, determinadas em laboratório, foram semeadas em caixas plásticas, medindo 45 cm x 30 cm x 11 cm, contendo aproximadamente 13 kg de solo previamente esterilizado. Para cada nível de incidência, foram semeadas 400 sementes a uma profundidade de 3 cm. O delineamento estatístico utilizado foi o de blocos completamente casualizados com seis tratamentos e quatro repetições por tratamento.
Sete dias após a sementeira foi feita a primeira avaliação, contando-se o número de plântulas emergidas com e sem sintomas de helmintosporiose. Foram consideradas as plântulas com sintomas na plúmula ou primeira folha. Aos 15 dias após a sementeira foi realizada nova avaliação. Decorridos 35 dias após a emergência, as plantas foram removidas e os coleóptilos e raízes foram lavados em água corrente e os sintomas examinados visualmente. No caso das raízes, para maior contraste no exame, estas foram emersas em água contida numa bandeja branca conforme Reis e Casa (1998).
A determinação da taxa de transmissão foi feita após a contagem de plântulas emergidas com sintomas, calculando-se a porcentagem em relação ao número de sementes emergidas, multiplicando-se esse valor por 100, dividindo-se pelo nível inicial de incidência do patógeno nas sementes, determinado em laboratório.
Análise de dados
A percentagem de germinação das sementes, as percentagens de transmissãode B. sorokiniana da semente para a plântula e planta bem como as percentagens obtidas nos parâmetros obtidos nos ensaios de campo depois de transformados em arcoseno raiz de X, foram submetidos a análise de variância. As médias obtidas foram comparadas pelo Teste de Duncan a 5% de probabilidade.
Para determinar a taxa de transmissão, após a contagem de plântulas emergidas com sintomas de helmintosporiose, calculou-se a porcentagem em relação ao número de sementes emergidas, multiplicando-se esse valor por 100, dividindo-se pelo nível inicial de incidência do patógeno nas sementes (Forcelini, 1991).
Os níveis de incidência do patógeno nas sementes foram correlacionados com a emergência e a porcentagem de plântulas infectadas. Os dados foram submetidos à análise de correlação linear aplicando-se o teste t.
Resultados e Discussão
Ensaio de campo
As médias de temperatura e pluviosidade registadas no período do ensaio de campo estão apresentados nas Figuras 1 e 2.
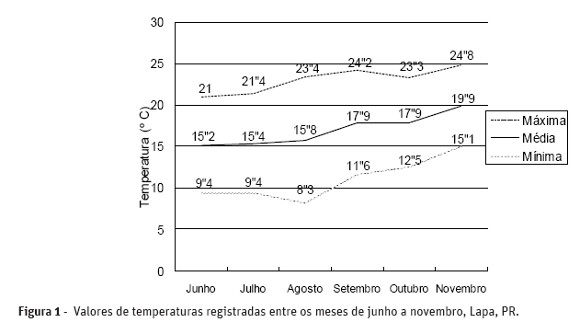
A intensidade da helmintosporiose variou de um ano para outro e de uma região para outra, provavelmente em função das variações de temperatura (Mehta et al., 1996). Pelos dados apresentados na Figura 1, constata-se que as temperaturas registradas no período de realização do estudo, foram suficientes para promover o estabelecimento da doença no campo, pois o fungo requer para germinação, infecção, crescimento da lesão e esporulação, temperatura oscilando entre 20 e 30 ºC (Luz e Bergstrom, 1986). Forcelini (1991) relata que para B. sorokiniana, o efeito da temperatura é dividido em três níveis: aos órgãos aéreos, a transmissão é maior na faixa térmica de 20-25 ºC, enquanto que 15-20 ºC restringe mais o patógeno ao sistema radicular e entre 25-30 ºC ocorre maior morte de plantas. A temperatura funcionaria como instrumento regulador da intensidade do processo. Como os conídios para germinarem e penetrarem nas folhas precisam de temperaturas entre 24 a 28 ºC e 9 a 24 horas de água livre sobre a superfície da folha por uma a duas semanas, as condições não foram propícias ao estabelecimento do patógeno no campo após o início do afilhamento (Quadro 1).
O estresse hídrico na floração afetou o desenvolvimento das cariopses, dado que provocou esterilidade floral principalmente na base e no ápice da espiga, diminuindo sensivelmente o número de carioses formados por espiga, produzindo cariopses pequenas e estéreis.
Durante o ensaio de campo, as temperaturas máximas oscilaram entre 21 e 24,8 ºC e as mínimas entre 9,4 a 15,1 ºC, favorecendo o desenvolvimento do patógeno no sistema radicular, o que talvez possa explicar a baixa emergência das plantas, observada no Quadro 2.
No entanto, quando se observa a Figura 2, nota-se uma baixa pluviosidade em todo o período, onde o total acumulado da sementeira até a colheita foi de 324 mm, que, em associação com a temperatura, promoveram condições para o desenvolvimento do patógeno na parte aérea desde a emergência das primeiras folhas.
O Quadro 2 mostra a transmissão de B. sorokiniana da semente para a planta sob condições de campo. Os níveis de infecção natural de B. sorokiniana nas amostras de sementes selecionadas em laboratório e usadas no ensaio de campo, variaram de 2,7% no tratamento 1, nível mais baixo até 55,5% no nível mais alto. Verificou-se a transmissão do fungo das sementes para a parte aérea, demonstrando dessa forma a capacidade do fungo de infectar as plantas no campo. O maior nível de incidência da doença em plantas no campo foi observado no tratamento 6 com 17%, seguido dos tratamentos 2, 4 e 5. Não foi significativa a correlação entre a incidência do fungo nas sementes, a emergência e a incidência nas plântulas. No tratamento 3 com nível de incidência de 12,5%, as plantas apresentaram a maior porcentagem de emergência (92%) e a mais baixa incidência do patógeno nas primeiras folhas.
No entanto, os resultados apresentados no Quadro 2 mostram também que houve grande variação na taxa de transmissão, concordando com os dados citados por Forcelini (1991) de que há pouca correlação entre o uso de sementes com níveis de infecção menores que 30%, por não causar a morte de plântulas, mas tem energia suficiente para permitir uma transmissão bem sucedida no campo. Essa hipótese explica o elevado potencial de plântulas emergidas, diferente do que se verificou para o tratamento 6, mais infectado, onde ocorreu uma transmissão elevada e emergência de apenas 70%. Provavelmente, o fungo presente na semente, durante o processo de geminação, infectou os coleóptilos, entrenós subcoronais e posteriormente as raízes seminais, levando à podridão das raízes que impediram a emergência das plantas.
A taxa de transmissão baseada nos sintomas foliares variou de 31% para o nível de 55,5% de incidência a 100% para os níveis de 2,5% e 7,5%, enquanto a relação semente infectada/planta doente aumentou na mesma medida que aumentou a incidência, variando de 0,3:1,0 no tratamento 1 e 3,2:1,0 no tratamento 6. Esses resultados confirmam aqueles encontrados anteriormente por Reis e Forcelini (1993) e Goulart (1996) para o patógeno na cultura do trigo.
No Quadro 3 são apresentados os dados de produtividade das parcelas, transformados em kg ha-1. Pode-se observar que o tratamento 1 com nível de incidência de 2,7, foi pouco afetado pela doença, visto que a infecção considerada elevada até os 14 dias após a emergência das plantas, se estabilizou, não havendo condições climáticas favoráveis para a transmissão à outras plantas.
A diferença de rendimento entre os tratamentos pode ser causada pela baixa emergência das plantas, devido à elevada incidência nas sementes, resultado também encontrado por Wiewióra (2006). Reis (1981) demonstrou que os sintomas de podridão de raízes seminais, bem como o de lesões nas primeiras folhas do trigo, podem ser causados por inóculo de B. sorokiniana presente na semente. Da semente, o inóculo foi trazido à superfície pelo coleóptilo e deste, à primeira folha.
Ensaios em estufa
Transmissão de B. sorokiniana da semente para a plântula: no Quadro 4, observa-se que a incidência do fungo nas primeiras folhas de plântulas de cevada foi inversamente proporcional à incidência nas sementes, isto é, quanto maior a incidência nas sementes, menor a porcentagem de infecção nas folhas. Para uma incidência de 2,5% nas sementes, verificou-se 6,2% de folhas infectadas, já para os níveis de 16,5%, 31,0% e 55,5% as folhas infectadas apresentaram 1,7%, 1,2% e 1,0%, respectivamente.
O fungo uma vez estabelecido na primeira folha poderia pela posterior esporulação e disseminação, estabelecer-se nos demais órgãos aéreos. A não manifestação dessa sintomatologia em outras plântulas e a não ocorrência de ciclos secundários nas demais folhas, leva à hipótese de que a semente infectada foi a fonte de inóculo primário, concordando com os dados obtidos por Reis (1981).
O fungo presente nas sementes provocou efeito negativo na emergência de plântulas, principalmente no tratamento 6 com apenas 76% de plântulas emergidas, uma vez que os dados de incidência inicial e nas raízes apresentam correlação positiva (r = 097*), mostrando que há correlação entre o aumento da incidência da doença nas raízes e a incidência do patógeno nas sementes. Os dados observados confirmam o trabalho conduzido por Reis (1981) de que o patógeno uma vez presente na semente ataca primeiro o coleóptilo e daí progride para a parte inferior, podendo matar a plântula antes da emergência.
Os valores de transmissão registrados para as primeiras folhas, coleóptilos e para raízes seminais são semelhantes, porém, diferentemente do que ocorre para as primeiras folhas, a incidência nas raízes e coleóptilos aumenta com a incidência do fungo nas sementes. Esses resultados diferem daqueles encontrados por Reis e Forcelini (1993), quando verificaram valores mais elevados de transmissão sintomática para coleóptilos, seguidos, em ordem decrescente, para raízes seminais e para plúmulas em trigo.
Transmissão de B. sorokiniana da semente para as primeiras folhas: no Quadro 5 são apresentados os resultados referentes à incidência de B. sorokiniana em sementes de cevada e sua relação com a transmissão do patógeno, para as primeiras folhas de plântulas.
Observou-se que a taxa de transmissão de B. sorokiniana diminuiu à medida que o nível de incidência nas sementes aumentou, porém existe grande diferença entre os tratamentos que segundo Forcelini (1991), quando se utilizam sementes com níveis de infecção menores que 30%, os resultados são controversos. Segundo o mesmo autor, existe pouca correlação com as análises de sanidade em laboratório, devido ao potencial de inóculo, que sendo menor não causa a morte de plântulas, mas é capaz de permitir uma transmissão bem sucedida a campo.
Com relação ao número de sementes infectadas/plantas doentes, o tratamento 1 apresentou a menor relação que foi de 0,4:1,0 e o tratamento 6 a maior, de 55,5:1,0, isto é, 55,5 sementes infectadas para 1 planta com sintoma na folha.
Transmissão de B. sorokiniana da semente para coleóptilos: no Quadro 6 apresentam-se os resultados referentes à incidência de B. sorokiniana em sementes de cevada e sua relação com a transmissão do patógeno, para os coleóptilos das plântulas. Observou-se que a transmissão do patógeno variou conforme a incidência nas sementes e a taxa de transmissão, baseada nos sintomas em coleóptilo aumentou proporcionalmente com o aumento da incidência nas sementes, sendo a relação semente infectada/ planta doente de 1,6 : 1 no tratamento 1; 4,6 : 1 no tratamento 5; e 10,6 : 1 no tratamento 6.
Transmissão de B. sorokiniana da semente para raízes: o Quadro 7 mostra os resultados referentes à incidência de B. sorokiniana em sementes de cevada e sua transmissão para as raízes seminais de plântulas. Verificou-se que há correlação entre a incidência do fungo nas sementes e a incidência em raízes seminais, pois houve um pequeno aumento da porcentagem de raízes infectadas com o aumento do nível de incidência. Os valores mais elevados de transmissão foram registrados para coleóptilos, seguidos para raízes seminais e para plúmulas. Observa-se ainda que a transmissão do patógeno variou conforme a incidência nas sementes e a taxa de transmissão. Com base nos sintomas em coleóptilo, infere-se que aumentou proporcionalmente com o aumento da incidência nas sementes, sendo a relação semente infectada/planta doente de 1,6:1 no tratamento 1; 4,6:1 no tratamento 5; e 10,6:1 no tratamento 6.
Houve decréscimo da taxa de transmissão para as raízes com o aumento da incidência do fungo nas sementes. Esses resultados reforçam a teoria de que o transporte do fungo pelas sementes não assegura, necessariamente, a sua transmissão às plantas no campo. Forcelini (1991) refere que em amostras de sementes onde foi feita desinfestação, as altas taxas de transmissão B. sorokiniana em trigo foram devidas à localização do patógeno no endosperma, sendo que a proporcionalidade entre o transporte e a transmissão foi observada apenas nos lotes com média e alta infecção.
Conclusões
Não houve correlação entre os diferentes níveis de incidência de B. sorokiniana nas sementes e a incidência em plantas no campo;
Houve correlação entre os diferentes níveis de incidência de B. sorokiniana nas sementes e a incidência em raízes primárias em plantas sob condições de estufa;
Níveis acima de 55% de B. sorokiniana nas sementes diminuiram drasticamente a emergência das plântulas, tanto em campo como em estufa.
Referências bibliográficas
Arias, G. (1995) - Mejoramiento genetico y produccion de cebada cervecera en America del Sur. Santiago, Chile: Direccion de produccion y proteccion Vegetal (FAO), Oficina Regional de la FAO para America Latina e Caribe, 162 p. [ Links ]
Barley (Hordeum) Maps and Statistics (2014) - Disponível em: http://www.gramene.org/species/hordeum/barley_maps_and_stats.html acesso em 09 junho de 2014. [ Links ]
Conab (2014) - Cevada Brasil: séries históricas. - Disponível em: http://www.conab.gov.br/OlalaCMS/uploads/arquivos/14_03_12_08_41_24_boletim_graos_marco_2014.pdf. Acesso em: 27 março 2014. [ Links ]
De Mori, C. e Minella, E. (2012) - Aspectos econômicos e conjunturais da cultura da cevada . Passo Fundo: Embrapa Trigo, 28 p. (Embrapa Trigo. Documentos Online, 139). Disponível em: http://www.cnpt.embrapa.br/biblio/do/p_do139.htm [ Links ]
Forcelini, C.A. (1991) - Importância epidemiológica de fungos do gênero Helminthosporium em sementes de trigo e cevada. In: Menten, J.O.M. (Ed.) Patógenos em sementes - detecção, Danos e Controle Químico. Piracicaba: ESALQ/FEALQ, p.179-190. [ Links ]
Goulart, A.C. (1996) - Transmissão de Bipolaris sorokiniana de sementes ao coleóptilo de trigo. Fitopatologia Brasileira, vol. 22, n. 1, p.05-08. [ Links ]
Lasca, C.C. (1997) - Padrões de sanidade e tolerância de infecção de sementes por patógenos de importância econômica. Summa Phytopathologica, vol. 23, n. 1, p. 85-86. [ Links ]
Luz, W.C e Bergstrom, G.C. (1986) - Temperature-sensitive development of spot blotch in spring wheat cultivars differing in resistance. Fitopatologia Brasileira, vol. 11, n. 1, p. 197-204. [ Links ]
Mehta, Y.R; Campos, L.A.C e Guzman, E. (1996) - Resistência genética de cultivares de trigo a Bipolaris sorokiniana. Fitopatologia Brasileira, vol. 21, n. 4, p. 455-459. [ Links ]
Neergard, P. (1979) - Seed Pathology. London, The MacMillan Press. England. 839p. [ Links ]
Picinini, E.C. e Fernandes, J.M. (1995) - Doenças em cereais de inverno - Aspectos epidemiológicos e controle, Passo Fundo: EMBRAPA-CNPT. 57 p. [ Links ]
Brasil (2009) - Regras para análise de sementes. Ministério da Agricultura, Pecuária e Abastecimento, Brasília, 2009, 399p. [ Links ]
Reis, E.M. (1981) - Podridão de raízes seminais e lesões foliares do trigo (Triticum aestivum L.) associados a Helminthosporium sativum Pam., King & Bakke, transmitido pela semente. Summa Phytopathologica, vol. 7, n. 3/4, p. 39-44. [ Links ]
Reis, E.M. e Casa, R.T. (1998) - Patologia de sementes de cereais de inverno. Passo Fundo: Aldeia Norte, 88p. [ Links ]
Reis, E.M. e Forcelini, C.A. (1993) - Transmissão de Bipolaris sorokiniana de sementes para órgãos radiculares e aéreos do trigo. Fitopatologia Brasileira. vol. 18, n. 1, p. 76-81. [ Links ]
Santos, H.P.; Reis, E.M.; Lhamby, J.C.B. e Sandini,I. (1995) - Características agronômicas e controle de doenças radiculares da cevada, em sistema plantio direto em rotação com outras culturas. Pesquisa Agropecuária Brasileira, vol. 30, n. 11, p. 1297-1303. [ Links ]
Wiewiora B. (2006) - Pathogenic ability of Bipolaris sorokiniana in relation to spring barley (Hordeum vulgare). Phytopathologia Polonica, vol. 41, p. 5-14. [ Links ]
Zadoks, JC.; Chang, TT e Konzak, CF. (1974) - A decimal code for the growth stages of cereals. Weed Research, vol. 14, n. 6, p. 415-421. [ Links ]
Recebido/received: 2014.06.12
Aceite/Accepted: 2015.04.17

