










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.38 no.2 Lisboa jun. 2015
ARTIGO
Relationships between leaf anatomical features of Arundo donax and glyphosate efficacy
Parâmetros morfoanatómicos foliares de Arundo donax e eficácia do glifosato
Ana Monteiro1*, Generosa Teixeira2 e Jorge Frazão Moreira3
1,* LEAF, Instituto Superior de Agronomia, Universidade de Lisboa, Tapada da Ajuda, 1349-017 Lisboa, Portugal. E-mail: anamonteiro@isa.ulisboa.pt, author for correspondence
2 Centre for Ecology, Evolution and Environmental Changes (CE3C), Faculdade de Farmácia, Universidade de Lisboa, Av. Prof. Gama Pinto, 1649-003 Lisboa, Portugal. E-mail: gteixeira@ff.ulisboa.pt
3 Direção Geral Agricultura e Veterinária, Quinta do Marquês, 2780-155 Oeiras, Portugal. E-mail: jfmoreira@dgadr.pt
ABSTRACT
Cuticle thickness is known to affect the amount of active ingredient that reaches the target site of a post emergence herbicide and consequently its efficacy. So, this work aimed to relate glyphosate efficacy with the thickness of leaf cuticle of young and mature leaves of Arundo donax. Glyphosate was applied at different development stages of the invasive species, in spring, summer and autumn at the concentrations of 0.675; 1.350 and 2.025 g L-1. Cuticle thickness of mature and young leaf blade and leaf sheath was measured on transverse sections by light microscopy. Despite the significant differences in the cuticle thickness between young and mature leaves, this did not seem affect the efficacy of glyphosate uptake since no significant differences were observed between young and mature leaves glyphosate treatments. But herbicide efficacy was dependent of the plant development stage. Higher efficacy of glyphosate was recorded when plots were sprayed at the end of October (flowering) compared to end April and mid June applications. Mortality of giant reed was higher than 90% one year after treatment only in autumn treatments. This result suggests that other mechanisms are involved, probably the reserves carrying over to rhizomes, on herbicide efficacy.
Key-words: Poaceae; invasive species; cuticule, leaf anatomy
RESUMO
É sabido que a espessura da cutícula afeta a quantidade dum herbicida de pós emergência que chega ao local de ação e consequentemente a sua eficácia. Assim, este trabalho teve por objetivo relacionar a eficácia do glifosato com a espessura da cutícula de folhas jovens e adultas da invasora ribeirinha Arundo donax. O glifosato foi aplicado em diferentes estágios de crescimento da invasora, na primavera, verão e outono nas concentrações de 0,675; 1,350 e 2,025 g L-1. A espessura da cutícula foi medida por recurso a microscopia ótica em cortes transversais de folhas adultas e jovens, limbo e bainha, de A. donax. Apesar de haver diferenças significativas na espessura da cutícula de folhas jovens e adultas da invasora, tal facto não afetou a eficácia do glifosato dado que não se registaram diferenças significativas nos tratamentos herbicidas quer nas plantas com folhas jovens quer nas com folhas adultas. Todavia a eficácia variou com o estado de crescimento. As eficácias mais elevadas e significativas foram observadas nos talhões tratados à floração, fim de outubro. Nestes, um ano após o tratamento químico a eficácia era superior a 90%. Estes resultados apontam para que outros mecanismos envolvidos na eficácia do glifosato, possivelmente devido a uma maior translocação para os órgãos de reserva, associada à translocação dos assimilados que ocorre neste período de crescimento.
Palavras-chave: Poaceae; invasoras; cutícula, anatomia da folha
Introduction
Arundo donax L., giant reed (cana, in Portuguese), is naturalized in Portugal and nowadays it is considered an invasive plant species. Another species Arundo plinii Turra is also found but only in the phytogeographic mainland Portugal regions of Center‑East and Center-South (Franco and Rocha-Afonso, 1998). Giant reed is a tall perennial cane found in riparian areas with well-drained soil, high temperatures and low salinity, forming dense stands on disturbed sites, sand dunes, in wetlands and riparian habitats. Its density increases in open places with high photosynthetic active radiation where native vegetation has been recently removed or damage.
A. donax generally grows to 6 m, in ideal conditions it can exceed 10 m, with hollow culms 2 to 3 cm diameter. The leaves are alternate, 30 to 60 cm long and 2 to 6 cm wide with a tapered tip, grey-green, and have a hairy tuft at the base Fig. 1A, B). Overall, it resembles an outsize common reed (Pragmites australis (Cav.) Steudel) but it is taller and more vigorous (Franco and Rocha-Afonso, 1998). A. donax flowers in late summer, bearing upright, feathery plumes 40 to 60 cm long, but the seeds are rarely fertile (Lewandowski et al., 2003). So, the reproduction is essentially asexual through vegetative propagation by underground vigorous rhizomes (Fig. 1C). The rhizomes are tough and fibrous and form knotty, spreading mats that penetrate deep into the soil (Mackenzie, 2004; Wijte et al., 2005; Mariani et al., 2010). The success of giant reed lies in part in their morphology and growth processes, and in part in their physiological diversity. Giant reed is a C3 plant, yet it displays the unsaturated photosynthetic potential of C4 plants, and is capable of extremely high photosynthetic rates (Rossa et al., 1998; Papazoglou et al., 2005), what gives the invasive species a high degree of effectiveness in the water use.

A waterside plant community dominated by A. donax may also reduce canopy shading of the in-stream habitat, which may result in increased water temperatures. This may lead to decreased oxygen concentrations and lower diversity of aquatic animals (Bell, 1997). The autochthones plant species diversity is also negatively affected. The culms and leaves have large amounts of toxic compounds like silica (Mackenzie, 2004), being an unpleasant food for insects and other animals (Miles et al., 1993).
Considering that A. donax affects the riparian structure (Herrera and Dudley, 2003), its ecological processes, namely the hydrological regime, erosion, sediments deposit and nutrients fluxes, this implies that its presence increases the risk of floods in the nearby areas. Its high water use and evapotranspiration also reduces the water availability and water stream. Abichandani (2007) estimated that the transpiration rate of giant reed could be 6 to 110 times higher than the one of the native species. Coffman et al. (2010) verified that after a fire giant reed density and biomass was 20 and 14-24 times, respectively, higher than the values observed for native plant species.
To remove giant reed, besides manual, there are mainly two methods, mechanical and/or chemical ones. Manual and mechanical control methods are very expensive and not effective since stem or rhizome pieces even containing a single node readily sprouted under a variety of conditions (Boose and Holt, 1999; Newhouser et al., 1999; Decruyenaere and Holt, 2001). Nevertheless these methods can be used any time (Oakins, 2001) being the best moment before plant flowering when the rhizomes reserves are low (Newhouser et al., 1999).
Giant reed chemical control trough ghyphosate foliar or stem applications is the mainly method used overall (Newhouser et al., 1999; Dudley (2005). Nevertheless, the uptake amount of foliar-applied herbicides usually varies with the morphological and anatomical traits of leaves, since an increase in the thickness of the cuticle layer may slower herbicide uptake (Wanamarta e Penner, 1989; Chachalis et al., 2001; Ferreira et al., 2002; Hoss et al., 2003; Huangfu et al., 2009), that is, the leaf surface and leaf anatomy may affect the efficacy of glyphosate uptake in weeds. Sherrick et al. (1986) investigated the effects of adjuvants and the environment during plant development on the level of glyphosate absorption and translocation in Convolvulus arvensis L. and confirmed that the thickness of the cuticle of weed affects the herbicide's level of efficacy.
For riparian plant species control, glyphosate was selected as the most appropriate product after specific considerations on efficacy, environmental safety, no soil residual activity, operator safety, application timing, and cost-effectiveness (Jackson, 1998). Single late-season applications of 1.350 g L-1 glyphosate onto the foliar mass showed efficacies varied between 50-95% one year after treatment (Newhouser et al., 1999). Previous stem cutting and thereafter glyphosate spraying 3-6 weeks later at the young leaves in order to reduce the amount of herbicide but to improve the spray distribution, was also recommended for large areas (Bell, 1997; Newhouser et al., 1999). Monteiro et al. (1999) observed that cutting common reed (P. australis) during flowering, in the year before herbicide treatment - doses of 2.9 and 3.24 kg a.i. ha-1 of glyphosate, applied at spring and at flowering -, improved the efficacy of the herbicide. Considering the botanical proximity between giant reed and common reed a similar response to glyphosate treatment could be expected.
In order to kill a perennial plant such as A. donax using systemic herbicides it is necessary to achieve a lethal dose in the perennial organs, in this instance the rhizomes of the plant. So, the main objective of this work was to maximize weed control using appropriate rates of glyphosate applied at various growth stages of the weed and their relationship to the plant leaf cuticle thickness.
Material and Methods
Morphological Studies
Leaves of A. donax were collected in South Portugal, Algarve, Silves, Algoz River. Samples of the blade and sheath median portions of mature and young leaves were studied by using light microscopy.
Light microscopy (LM): Small pieces of plant material were fixed in a 3% glutaraldehyde solution for 5 h at 4°C, followed by 0.1 M phosphate buffer wash, pH 7.0, during 24-h at 4 °C; three distilled water washes, of 30-min each and dehydrated in a graded series ethanol solutions (30%, 50%, 70% and 100%, 20 to 40-min each) (Hayat, 2000). Some fixed material was embedded in paraffin (Ruzin, 1999) and tissue blocks were sectioned at 10-12 μm using a Leitz 1512 Minot microtome. Transverse sections could be clarified in Chloral Hydrate, stained with acidic Phloroglucin to highlight lignified plant tissues or instead with Sudan Red for total lipids (Johansen, 1940) to highlight the cuticle. Observations were carried out under a Nikon Eclipse E400 microscope equipped with a Nikon Coolpix MDC lens adapter. Images were obtained with a Nikon Coolpix 995 digital camera. Qualitative and quantitative characters are the average of 20 different observations.
Chemical control studies
Dense infestations of A. donax - 16 to 27 shoots per m-2 - growing in Algarve, Portugal, Silves, Algoz River, were used for the study. A. donax plants were cut one month before spring and summer, and one year before autumn herbicide applications. Application dates were at end April, middle June and end October 2010.
The cut plots were cleared of vegetation to within 15 cm of the surface using a hand-held weed trimmer equipped with a saw blade. Glyphosate concentrations of 0.675, 1.350 and 2.015 g L-1 (Roundup® Supra, 450 a.i. g L-1) were applied at 600 L ha-1. Applications were made using a hand held lance with an adjustable nozzle. The medium output, at the pressure of 10 bars, was 1.8 L/min. Plots 7 x 20 m were arranged end-to-end on the bank of the river in a split-plot design with three replications. Plant cutting was the main factor, glyphosate concentrations the subplot factor. Stand reduction was measured as percentage cover by three independent observers, 12 months after treatment. Percent A. donax control was rated visually, based on a scale of 0-100 (0 = no reduction in weed biomass and 100 = no living weeds present).
Data analysis
Cuticle thickness values were subjected to analysis of variance and separated using Fisher's Protected LSD test at the 5% level of significance.
Estimates of A. donax cover percentages were averaged before the data were subjected to analysis of variance. Cover percentages were subjected to angular transformation before analysis of variance. Means of percentage cover (transformed) were separated using Fisher's Protected LSD test at the 5% level of significance.
Results and discussion
Leaf anatomy
A. donax leaf blades cross sections show a typical monocot organization. The dermal system is represented by the epidermis of the inner face and the outer face. Between these the mesophyll is undifferentiated and the veins all appear parallel to each other. The vascular bundles are surrounded by two bundles sheaths, an inner one called the mestome sheath that shows thick walled cells, which also stained positive with acidic Phloroglucin. The mestome sheath is encircled by an outer sheath of non-photosynthetic larger thin walled cells which are surrounded by mesophyll cells (Fig. 2A). This is the pattern anatomy characteristic in Poaceae and related with C4 photosynthesis, the called Kranz anatomy (Esau, 1997).
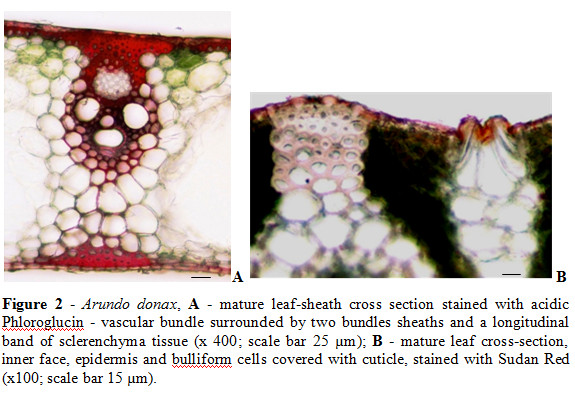
Associated with the vascular bundles, in addition to the double sheath, there are longitudinal bands of sclerenchyma tissue (Fig. 2A). In mature leaf blades and in their leaf sheaths there is also a continuous band of sclerenchyma throughout the epidermis or, in the absence of this, involving the hypodermal cells. In young leaf blades and in their leaf sheaths there is no continuous band below the protection tissue but there are already cells with thickening in tangential cell walls.
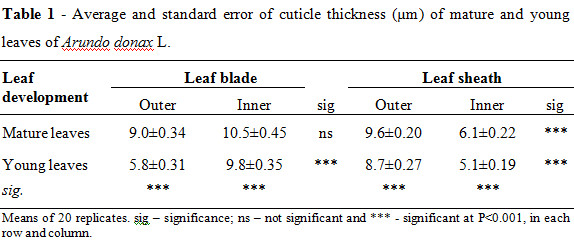
In A. donax leaf blades and leaf sheaths cross sections it is also visible the presence of the cuticle, covering both faces epidermal tissues. Table 1 shows the cuticle thickness of A. donax mature and young leaf blades and leaf sheaths, external and internal surfaces. Significantly cuticle thickness differences were observed between mature and young blade and sheath leaves. The outer and inner cuticles, with exception for mature blade leaves, revealed also significant differences on their thickness.
The epidermis of the inner face can also include, besides the stomata, other enlarged cell types designated as motor or bulliform cells, also covered with a thick cuticle (Fig. 2B). These cells are associated with the winding of the leaves in dry conditions.
Glyposate efficacy
One year after treatment, the efficiency of glyphosate over A. donax significantly increases with its concentration (Table 2). Concentrations below 1.350 g a.i. L-1 of glyphosate were inefficient in the chemical control of A. donax applied in young (cut plots) or adult plants (uncut plots), at any application date. But glyphosate efficacy was significantly increased when applied at giant reed flowering both in cut and uncut plots.
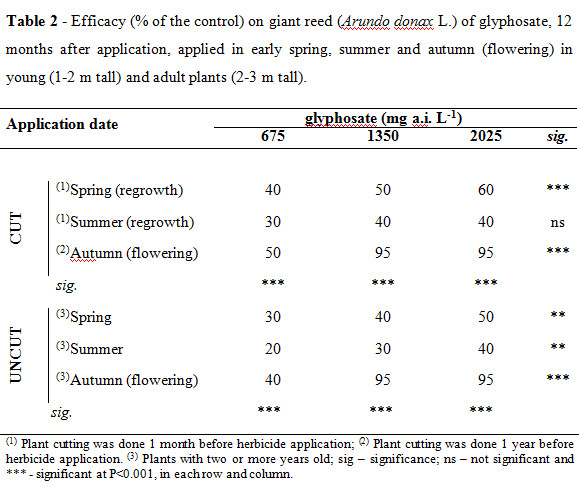
The lowest glyphosate efficacies were observed for all summer treatments. That is, higher efficacy of glyphosate was recorded when plots were sprayed at the end of October (flowering) compared to end April and mid June applications. Mortality of giant reed was higher than 95% one year after treatment only in autumn treatments, probably because during this phenological stage reserves are being carrying over to the rhizomes (Bell, 1997; Decruyenaere and Holt, 2001). These results were similar to the ones obtained by several authors (Newhouser et al., 1999; Spencer et al., 2008) for giant reed where the spring and summer glyphosate treatments do not seem appropriate, but contrary to those obtained by Monteiro et al. (1999) for common reed,
According to several authors, the uptake amount of foliar-applied herbicides usually varies with the morphological and anatomical traits of leaves, since an increase in the thickness of the cuticle layer may slower herbicide uptake (Wanamarta e Penner, 1989; Chachalis et al., 2001; Ferreira et al., 2002; Hoss et al., 2003; Huangfu et al., 2009), that is, the leaf surface and leaf anatomy may affect the efficacy of glyphosate uptake in weeds. Sherrick et al. (1986) investigated the effects of adjuvants and the environment during plant development on the level of glyphosate absorption and translocation in Convolvulus arvensis L. and confirmed that the thickness of the cuticle of weed affects the herbicide's level of efficacy. Our study clearly illustrates morphological disparities in the epidermal and anatomical features of young and mature leaves of A. donax, but the differences did not explain their susceptibility difference to glyphosate.
Furthermore, there might be other mechanisms that are responsible for herbicide response differences, namely differences in the translocation patterns that might affect the efficacy of glyphosate in giant reed (Sandberg et al., 1980; Monquero et al., 2005). Glyphosate is amphimobile in plants and is highly translocate from the site of application through the phloem and xylem to shoot and root sinks tissues, following the plant carbon flow (Dewey and Appleby, 1983; Preston and Wakelin, 2008). This behaviour helps to explain the good activity of the herbicide at post flowering since, according to Soetaert et al. (2004) with common reed, at the onset of senescence, assimilates are allocated to belowground organs, that is the rhizomes. That behaviour probably explain giant reed response to glyphosate treatments.
Conclusions
Overall, the present study did not give support to the concept that the morphological and anatomical characteristics, in particular the cuticle thickness, of giant reed leaf influence the behaviour of glyphosate. Indeed, these results suggest that the translocation of the reserves to the rhizomes during and pos- flowering could be the force that increases glyphosate activity. The discrepancy observed in spring and summer treatments is likely due to the difference in the time of year when the plant grown is affected by other unknown factors, probably an higher herbicide translocation to rhizomes associated with the assimilate translocation during the blooming season could explain the results.
The results also pointed out that for an effective giant reed control more than one glyphosate application (1.350 g L-1) is necessary and the previous stem cutting is recommended in order to reduce the amount of herbicide and to improve the spray distribution.
Acknowledgments
The chemical studies were supported by the RICOVER Project SOE1/P2/P248 “River Recuperation in European SUDOE”.
References
Abichandani, S.L. (2007) - The potential impact of the invasive species Arundo donax on water resources along the Santa Clara River: Seasonal and Diurnal Transpiration. Master's thesis. Environmental Health Sciences, University of California, Los Angeles. 44 p. [ Links ]
Bell, G.P. (1997) - Ecology and management of Arundo donax, and approaches to riparian habitat restoration in Southern California. In: J.H. Brock, M. Wade, P. Pysek and D. Green, (Eds.) - Plant Invasion: Studies from North America and Europe. Leiden, the Netherlands, p. 103-113. [ Links ]
Boose, A.B. and Holt, J.S. (1999) - Environmental effects on asexual reproduction in Arundo donax. Weed Research, vol. 39, n. 2, p. 117-127. [ Links ]
Chachalis D., Reddy, K.N. and Elmore, C.D. (2001) - Characterization of leaf surface, wax composition, and control of redvine and trumpet creeper with glyphosate. Weed Science, vol. 49, n. 2, p. 156–163. [ Links ]
Coffman, G.C., Ambrose R.F. and Rundel, P.W. (2010) -Wildfire promotes dominance of invasive giant reed (Arundo donax) in riparian ecosystems. Biological Invasions, vol. 12, n. 8, p. 2723-2734. [ Links ]
Decruyenaere, J.G. and Holt, J.S. (2001) - Seasonality of clonal propagation in giant reed. Weed Science, vol. 49, n. 6, p. 760-767. [ Links ]
Dudley, T. (2005) - Global Invasive Species Database: Arundo donax. Invasive Species Specialist Group (ISSG) of the World Conservation Union. Available online at 2014.12.18: http://www.issg.org/database/species/ecology.asp?si=112 [ Links ]
Dewey S.A. and Appleby A.P. (1983) - A comparison between glyphosate and assimilate translocation patterns in tall morning glory (Ipomoea purpurea). Weed Science, vol. 31, n. 3, p. 308-314. [ Links ]
Esau, K. (1997) - Anatomia das Plantas com Sementes. Editora Edgard Blucher, Ltda, S. Paulo, Brasil. 293 p. [ Links ]
Ferreira, E.A.; Procópio, S.O.; Silva, E.A.M.; Silva, A.A. and Rufino, R.J.N. (2002) - Estudos anatômicos de folhas de plantas daninhas. I - Nicandra physaloides, Solanum viarum, Solanum americanum e Raphanus raphanistrum. Planta Daninha, vol. 20, n. 2, p.159-167 [ Links ]
Franco, J.A. and Rocha-Afonso, M.L. (1998) - Nova Flora de Portugal (Continente e Açores). Vol. III (Fascículo II). Gramineae. Escolar Editora. Lisboa. 283 p. [ Links ]
Hayat M. (2000) - Principles and techniques of electron microscopy: biological applications. 4 Ed., Cambridge University Press. 543 p. [ Links ]
Herrera, A. M. and Dudley, T.L. (2003) - Invertebrate community reduction in response to Arundo donax invasion at Sonoma Creek. Biological Invasions, vol. 5, n. 1, p. 167-177. [ Links ]
Hoss, N.E.; Khatib, K.; Peterson D.E. and Loughin, T.M. (2003) - Efficacy of glyphosate, glufosinate, and imazethapyr on selected weed species. Weed Science, vol. 51, n. 1, p. 110–117. [ Links ]
Huangfu, C.; Song, X. and Qiang, S. (2009) - Morphological disparities in the epidermal and anatomical features of the leaf among wild Brassica juncea populations. Weed Biology and Management, vol. 9, n. 3, p. 234–242. [ Links ]
Jackson, N.E. (1998) - Chemical control of giant reed (Arundo donax) and saltcedar (Tamarix ramosissima). In: Bell, Carl E., (Ed.) - Arundo and saltcedar: the deadly duo. Proceedings of a workshop on combating the threat from arundo and saltcedar. 1998 June 17; Ontario, CA. Holtville, CA: University of California, Cooperative Extension: p. 33-42. [ Links ]
Johansen, D.A. (1940) - Plant microtechnique. New York: McGraw-Hill. 523 p. [ Links ]
Lewandowski, I. J.; Scurlock, M.O.; Lindvall, E. and Christou, M. (2003) - The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe. Biomass and Bioenergy, vol. 25, n. 4, p. 335-361. [ Links ]
Mackenzie, A. (2004) - Giant Reed. In: C. Harrington e A. Hayes (eds) - The Weed Workers' Handbook. The Watershed Project and California Invasive Plant Council, California. p. 92-93. [ Links ]
Mariani, C.; Cabrini, R.; Danin, A.; Piffanelli, P.; Fricano, A.; Gomarasca, S.; Dicandilo, M.; Grassi F. and Soave, C. (2010) Origin, diffusion and reproduction of the giant reed (Arundo donax L.) a promising weedy energy crop. Annals of Applied Biology, vol. 157, n. 2, p. 191–202. [ Links ]
Miles, D.H.; Tunsuwan, K.; Chittawong, V.; Kokpol, U.; Choudhary M.I. and Clardy J. (1993) - Boll weevil antifeedants from Arundo donax. Phytochemistry, vol. 34, n. 2, p. 1277-1279. [ Links ]
Monquero, P.A.; Cury, J.C. and Christofoletti, P.J. (2005) - Controle pelo glyphosate e caracterização geral da superfície foliar de Commelina benghalensis, Ipomoea hederifolia, Richardia brasiliensis e Galinsoga parviflora. Planta daninha, vol. 23, n. 1, p. 123-132. [ Links ]
Monteiro, A.; Moreira I. and Sousa, E. (1999) - Effect of prior common reed (Phragmites australis) cutting in herbicide efficacy. Hydrobiologia, vol. 415, n. 0, p. 305-308. [ Links ]
Newhouser, M.; Cornwall C. and Dale R. (1999) - Arundo: A Landowner Handbook. Available online at 2014.12.18: http://teamarundo.org/education/landowner_handbook.pdf [ Links ]
Oakins, A.J. (2001) - An Assessment and Management Protocol for Arundo donax in the Salinas Valley Watershed. Monterey Bay, CA: California State University. 51 p. [ Links ]
Papazoglou, E.G.; Karantounias, G.A.; Vemmos S.N. and Bouranis, D.L. (2005) - Photosynthesis and growth responses of giant reed (Arundo donax L.) to the heavy metals Cd and Ni. Environment International, vol. 31, n. 2, p. 243-249. [ Links ]
Preston, C. and Wakelin, A.M. (2008) - Resistance to glyphosate from altered herbicide translocation patterns. Pest Management Science, vol. 64, n. 4, p. 372-376. [ Links ]
Rossa, B.; TuAers A.V.; Naidoo G. and Willert, von D. J. (1998) - Arundo donax L. (Poaceae) — a C3 species with unusually high photosynthetic capacity. Botanica Acta, vol. 111, n. 3, p. 216–221. [ Links ]
Ruzin, S. (1999) - Plant microtechnique and microscopy. London: Oxford University Press. 322 p. [ Links ]
Sandberg, C.L.; Meggitt, W.F. and Penner, D. (1980) - Absorption, translocation and metabolism of 14C-glyphosate in several weed species. Weed Research, vol. 20, n. 4, p. 195-200. [ Links ]
Soetaert, K.; Hoffmann, M.; Meire, P.; Starink, M.; van Oevelen, D.; Van Regenmortel, S. and Cox, T. (2004) - Modeling growth and carbon allocation in two reed beds (Phragmites australis) in the Scheldt estuary (Belgium, The Netherlands). Aquatic Botany, vol 79, n. 3, p. 211-234. [ Links ]
Spencer, D.F.; Tan, W.; Liow, P.; Ksander, G.; Whitehand, L.C.; Weaver, S.; Olson J. and Newhauser, M. (2008) - Evaluation of glyphosate for managing giant reed (Arundo donax). Invasive Plant Science Management, vol. 1, n. 3, p. 248–254. [ Links ]
Sherrick, S.T.; Holt, H.A. and Hess, F.D. (1986) - Effects of adjuvants and environment during plant development on glyphosate absorption and translocation in field bindweed (Convulvus arvensis). Weed Science, vol. 34, n. 6, p. 811-816. [ Links ]
Wanamarta, G. and Penner D. (1989) - Foliar absorption of herbicides. Reviews of Weed Science, vol. 4, p. 215‑231. [ Links ]
Wijte, A.H.B.M.; Mizutani, T.; Motamed, E.R.; Merryfield, M.L.; Miller, D.E. and Alexander, D.E. (2005) - Temperature and endogenous factors cause seasonal patterns in rooting by stem fragments of the invasive giant reed, Arundo donax (Poaceae). International Journal of Plant Science, vol. 166, n. 3, p. 507-517. [ Links ]
Received/Recebido: 2014.12.08
Accepted/Aceite: 2015.02.09

