










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.37 no.4 Lisboa dez. 2014
ARTIGO
Suplementação dietética de fitase na metabolização de nutrientes nas fases inicial e crescimento de frangos de corte em estresse por calor
Dietary phytase supplementation on nutrient metabolism in initial and growth stages of broilers under heat stress
Domingos U. Carvalho Filho1, Agustinho V. Figueirêdo, João B. Lopes, Daniela C. P. Lima, Snaylla N. O. Almendra, Pedro E. Bitencourt e Vânia B. S. Lima
1 Departamento de Zootecnia, Centro de Ciências Agrárias ,Universidade Federal do Piauí (UFPI), Bairro Ininga, Teresina, PI, Brasil. E-mails: urquizafilho@hotmail.com, author for correspondence; agustinhov@yahoo.com.br; lopesjb@uol.com.br; danicrisvet@hotmail.com; snaylla@gmail.com; pedropiripiri@hotmail.com; vania_vet06@hotmail.com
RESUMO
Avaliou-se o efeito da inclusão de diferentes níveis de fitase nas dietas de frangos de corte sob estresse por calor nas fases inicial e crescimento nas variáveis: energia metabolizável aparente corrigida para o balanço de nitrogénio, coeficiente de metabolizabilidade da matéria seca, consumo, excreção e coeficiente de retenção aparente do nitrogénio, do cálcio e do fósforo. As aves foram distribuídas em delineamento de blocos casualizados e tratadas com cinco níveis de fitase (0, 1000, 2000, 3000 e 4000 FTU/kg da dieta) e cinco repetições. Os frangos receberam dietas isonutritivas à base de milho e farelo de soja, formuladas para atender as exigências nutricionais, exceto para fósforo disponível, que foi reduzido em 20% da exigência. Cada período experimental teve a duração de oito dias, em que quatro eram para adaptação das aves às gaiolas e às dietas e quatro para a Coleita das excretas. As dietas suplementadas com fitase reduziram a energia metabolizável aparente corrigida para o balanço de nitrogénio e diminuíram a excreção de fósforo das aves sob estresse por calor, nas fases inicial e crescimento.
Palavras-chave: estresse por calor, fitato, fitase, fósforo, frangos de corte
ABSTRACT
The experiments were conducted to evaluate the effect of different inclusion levels of phytase in diets on the apparent metabolizable energy corrected for nitrogen balance, metabolization coefficient of dry matter, intake, excretion and coefficient of apparent retention of nitrogen, calcium and phosphorus in broilers under heat stress in initial and growth stages. Birds were distributed in a randomized block design and treated with five levels of phytase (0, 1000, 2000, 3000 and 4000 FTU/kg diet) and five replicates. Isonutrient diets based on corn and soybean meal, fed on chickens were formulated to meet nutritional requirements, except for available phosphorus, which was reduced in 20 % of requirement. Each experimental period lasted eight days, four for adaptation of poultry cages and diets and four for collection of excreta. Diet supplemented with phytase reduced apparent metabolizable energy corrected for nitrogen balance and decrease phosphorus excretion of birds under heat stress, initial and growth stages.
Keywords: broiler, heat stress, phosphorus, phytase, phytate
Introdução
Nas formulações das dietas para frangos de corte, os ingredientes mais utilizados são milho e farelo de soja, e aproximadamente 66% do fósforo destes grãos estão na forma de ácido fítico, molécula orgânica que as plantas utilizam para armazenar fósforo, e que o torna indisponível para monogástricos (Buso et al., 2011).
A ligação do ácido fítico a um nutriente (hidrato de carbono, proteína ou mineral), no trato digestivo dos animais não ruminantes, dá origem ao sal denominado fitato. Esta molécula é um potente agente quelante de cálcio, cobre, zinco, magnésio, manganês e ferro, complexa-se também com proteínas, aminoácidos e hidrato de carbonos, além de inibir a atividade de algumas enzimas digestivas como: pepsina, tripsina e alfa-amilase (Lelis et al., 2010). Os complexos formados são insolúveis e prejudicam a disponibilidade dos nutrientes, com reflexo nas variáveis de desempenho dos animais monogástricos (Teixeira et al., 2013).
A incorporação de fósforo inorgânico à dieta das aves é um recurso nutricional utilizado devido à reduzida disponibilidade deste mineral nos ingredientes de origem vegetal. Entretanto, esta suplemento pode elevar os teores de fósforo nas excretas (Laurentiz et al., 2009; Gomide et al., 2011), e se não for dado destino adequado à cama, pode ocorrer elevação excessiva deste mineral nos mananciais de água e provocar eutrofização (Lelis et al., 2009).
O organismo das aves não produz fitase, todavia, o uso de fitase exógena na dieta tem sido eficiente em hidrolizar o fitato e disponibilizar fósforo, cálcio e outros minerais, além de melhorar a eficiência de utilização de proteínas/aminoácidos e energia das dietas das aves (Selle e Ravindran, 2007; Fukayama et al., 2008). Perante isso, diversos autores, dentre eles, Silva et al. (2006), Laurentiz et al. (2009), Gomide et al. (2011) e Silva et al. (2012) têm demonstrado que dietas com redução de fósforo disponível (Pd) e proteína bruta (PB), suplementadas com fitase, têm proporcionado redução do fósforo e nitrogénio nas excretas das aves, sem perda de desempenho.
Diversos investigadores relataram o efeito da fitase aumentando a disponibilidade e retenção do fósforo fítico de ingredientes vegetais em aves (Silva et al., 2008; Lelis et al., 2010; Gomide et al., 2011; Meneghetti et al., 2011). Entretanto, no que diz respeito à atividade da fitase sobre a disponibilidade de outros minerais, são encontrados dados contraditórios, indicando a necessidade de mais investigação sobre o assunto.
Mitchel e Edwars Jr. (1996) observaram que para uma maior solubilização do fitato no trato digestivo das aves é preciso manter os níveis de cálcio (Ca) e fósforo (P) nos limites mínimos necessários. A alta concentração de Ca ou a elevada relação Ca:Pd na dieta eleva o pH intestinal, reduz a fração solúvel dos minerais e sua disponibilidade para absorção, mesmo em presença de fitase.
Outro aspecto importante a considerar é a origem e o nível da enzima a ser adicionado. O nível da enzima deve estar de acordo com o nível da redução nutricional estabelecida, e do teor de fósforo complexado no ingrediente. A ação da fitase é dose dependente, e aumenta à medida que níveis mais altos são adicionados à dieta. Em contrapartida, à medida que os níveis nutricionais são reduzidos abaixo da exigência do animal, ocorre naturalmente, uma maior retenção do nutriente pelo organismo, para que sejam mantidas as funções fisiológicas, e neste caso, níveis mais altos de fitase podem ser necessários para que seja expresso o efeito da enzima (Shoener et al., 1993).
A temperatura da pele das aves em ambiente termoneutro (30 a 20 oC, a depender da idade, e umidade relativa do ar de 60 a 70%) mantém-se em torno de 33 oC, e a temperatura interna do corpo por volta de 41 oC (Welker et al., 2008, Aviagen, 2009). A diferença de temperatura entre a pele e o ambiente promoverá um fluxo de calor, neste caso, da pele (mais quente) para ambiente (mais frio). Este processo de transferência de calor dá-se por mecanismos radiativos, condutivos ou convectivos, com pouco dispêndio de energia pelas aves.
Contudo, quando a diferença de temperatura entre a pele e o ambiente se aproxima de zero, dificulta a perda de calor por esses mecanismos sensíveis, e ocorre aumento da temperatura interna das aves. Nessas circunstâncias, ativa-se o processo evaporativo de perda de calor (taquipnéia), entretanto, esse mecanismo de dissipação de calor tem alto custo energético, com reflexo negativo nos parâmetros produtivos das aves, podendo ainda alterar o pH sanguíneo (alcalose respiratória), e elevar a mortalidade da criação (Oliveira et al., 2006a; Oliveira et al., 2006b).
Aves em estresse por calor reduzem o consumo de ração na tentativa de minimizar a produção de calor gerado pelos processos de digestão e absorção de nutrientes, e a suplementação de fitase pode ser uma alternativa para aumentar a densidade nutricional relativa das dietas, uma vez que disponibiliza mais energia, aminoácidos e minerais das dietas no trato digestivo das aves, com possibilidade de minimizar as perdas de desempenho devido à redução do consumo.
Na ausência de informações da ação da fitase sobre o fitato, nas aves em condições de estresse por calor, conduziu-se este trabalho com o objetivo de avaliar o efeito da inclusão de diferentes níveis de fitase nas dietas, sobre as variáveis: energia metabolizável aparente corrigida para o balanço de nitrogénio, coeficiente de metabolizabilidade da matéria seca, consumo, excreção e coeficiente de retenção aparente do nitrogénio, do cálcio e do fósforo, em frangos de corte em estresse por calor, nas fases inicial e crescimento.
Material e Métodos
O estudo foi conduzido nos meses de agosto e setembro, no Setor de Avicultura do Departamento de Zootecnia do Centro de Ciências Agrárias da Universidade Federal do Piauí, no município de Teresina, estado do Piauí, Brasil, cujas coordenadas geográficas são: latitude de 5° 5' sul, longitude de 42° 48' oeste, altitude de 74,4 metros e clima tropical semi-úmido, com precipitação pluviométrica anual em torno de 1396 mm (Feitosa, 2010).
Foram realizados dois ensaios de metabolismo, ambos, com frangos machos da linhagem Ross. O ensaio para a fase inicial (ensaio I) foi realizado com 125 aves de 13 dias de idade, com 263±14,57 g em média; e o da fase de crescimento (ensaio II) foi conduzido com 100 aves de 22 dias de idade, e peso médio de 850±47,87 g. Todas as aves, antes do período experimental, foram mantidas em dietas a base de milho e farelo de soja, que atendiam às exigências segundo Rostagno et al. (2011). Posteriormente foram alojadas em gaiolas metabólicas preparadas com comedouros e bebedouros tipo calha e bandejas coletoras de excretas.
As aves foram distribuídas em delineamento de blocos casualizados (retirado a influência do peso) e submetidas a cinco tratamentos com cinco repetições. Os tratamentos foram caracterizados por diferentes níveis de fitase [Ronozyme NP (CT)]: 0; 1000; 2000, 3000 e 4000 FTU/kg de ração farelada, incluída em substituição de parte do material inerte (caulim). As unidades experimentais continham cinco (ensaio I) e quatro (ensaio II) aves, totalizando 25 parcelas em cada ensaio.
Cada período experimental teve duração de oito dias, sendo os quatro primeiros para adaptação das aves às gaiolas e às dietas, e os quatro últimos dias para a coleta das excretas. As dietas (Quadro 1) de cada fase tinham as mesmas composições centesimais e foram formuladas segundo recomendações preconizadas por Rostagno et al. (2011), exceto para fósforo disponível, que foi reduzido em 20%, tendo composição centesimal de 0,312% de Pd para a fase inicial e 0,273% de Pd para a fase de crescimento.
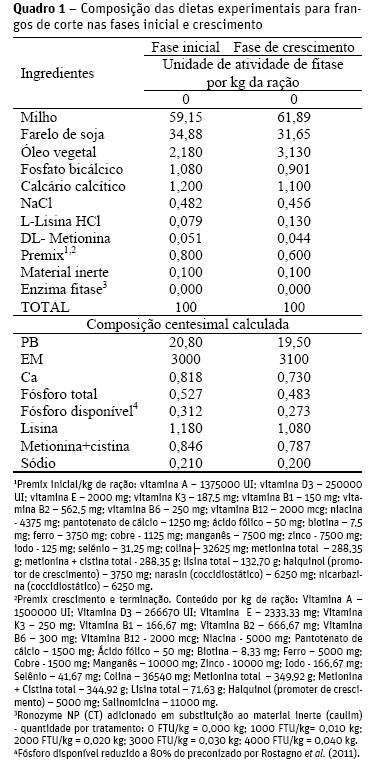
Como a concentração de cálcio não foi reduzida, as relações dietéticas Ca:Pd nas duas fases foram de 2,62:1 e 2,67:1, respectivamente.
A monitorização da temperatura e umidade relativa do ar no interior do galpão foi feita por recurso a termohigrómetro, colocado à altura intermediária das gaiolas. As leituras foram realizadas diariamente às 8:30-h e 15:30-h.
Durante o período experimental, a água foi fornecida ad libitum e trocada duas vezes ao dia para evitar aquecimento. O consumo de ração também foi ad libitum para os dois ensaios, sendo os comedouros supridos de ração duas vezes ao dia para evitar desperdícios. O programa de luz foi contínuo (natural e artificial), durante as 24 horas do dia.
Em cada ensaio, foram realizadas duas colheitas totais diárias das excretas (8-h e 14-h). As excretas foram acondicionadas em sacos plásticos devidamente identificados, pesados e armazenados em freezer a -18 °C, para posterior realização das análises laboratoriais.
As excretas provenientes das mesmas unidades experimentais, de cada ensaio, foram descongeladas e misturadas uniformemente. Depois foram pré-secas em estufa com ventilação forçada durante 72-h a 65°C. Em seguida, foram moídas e acondicionadas em recipientes plásticos para posteriormente, no laboratório de análise de alimentos do Departamento de Zootecnia da Universidade Federal do Piauí, ser analisado o teor de matéria seca (MS), energia bruta (EB) e nitrogénio (N), e no laboratório de bromatologia da Embrapa Meio Norte ser determinado nível de cálcio e de fósforo. As rações e excretas foram analisadas segundo metodologia descrita por Silva e Queiroz (2002).
No final dos ensaios foi determinada a quantidade de ração consumida por unidade experimental no período da coleta, e determinados os valores da energia metabolizável aparente corrigida para o balanço de nitrogénio (EMAn), coeficiente de metabolizabilidade da matéria seca (CMMS), coeficiente de retenção aparente do nitrogénio (CRN), do cálcio (CRCa) e do fósforo (CRP), utilizando-se as equações: EMAn (kcal/kg) = [EB consumida - EB das excretas - (BN x 8,22)] / MS ingerida; CMMS (%) = [(MS consumida – MS excretada) / MS consumida] x 100 e CR do mineral (%) = [(mineral ingerido - mineral excretado) / mineral ingerido] x 100, conforme Santos et al. (2011) e Gomide et al. (2011). Os dados foram submetidos à análise de variância e regressão polinomial, segundo o procedimento GLM do Statistical Analysis System – SAS (1996).
Resultados e Discussão
Os valores de temperatura e umidade relativa do ar registrados no interior do galpão, no período experimental foram, respectivamente, 28,5 ± 2,7ºC e 58,1 ± 15,62% durante o ensaio I, e 28,2 ± 1,5ºC e 53,1 ± 12,91% no ensaio II.
O ambiente de conforto é caracterizado quando o calor proveniente do metabolismo é dissipado para o ambiente sem prejuízo no desempenho. Esta situação de conforto é evidenciada, a partir da 2ª semana de vida, em locais com temperatura de 18 a 23 °C e umidade relativa do ar de 50 a 70% (Cella et al., 2001; Oliveira et al., 2006a; Oliveira et al., 2006b). Isso demonstra que estas aves, principalmente as da fase de crescimento, foram expostas a desconforto térmico. Temperatura acima da zona termoneutra, sobretudo com umidade relativa do ar acima de 70%, dificulta a dissipação de calor das aves por processo evaporativo, e provoca mudanças no comportamento e alterações metabólicas, com consequente queda do desempenho das aves (Oliveira et al., 2006a; Oliveira et al., 2006b; Aviagen, 2009).
Observou-se redução linear da EMAn em função dos níveis de fitase na fase inicial e na fase de crescimento (Quadro 2), segundo as equações: y = 3369,262276 – 0,026017x (R2 = 0,679; P<0,05) e y = 3501,83 – 0,025526x (R2 = 0,595; P<0,01), respectivamente. Estas reduções na EMAn podem estar associados a ineficiência enzimática sobre os substratos energéticos complexados, devido as dietas atenderem as exigências em energia das aves, visto que Meneghetti et al. (2011) ao avaliarem dietas suplementadas com fitase (1500, 3000, 4500, 6000, 8000 e 10000 FTU/kg), com redução na energia metabolizável de 85 (2915 kcal/kg) e 60 (3040 kcal/kg) kcal/kg, respectivamente para as fases inicial e crescimento, afirmaram que as aves recuperaram a redução da energia dietética pelo aumento da digestibilidade do amido (hidrolise do complexo fitato-hidrato de carbono/fitase-amilase), e/ou por efeito indireto da suplementação enzimática, que minimizou reações de saponificação no trato digestivo entre os lipídios da dieta e os minerais do complexo fitato-mineral.
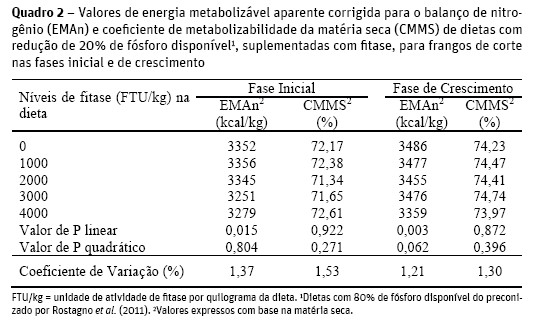
Santos et al. (2011) atendendo às exigências em energia das aves, observaram que rações suplementadas com fitase (500 FTU/kg), contendo 0,36% de Pd, também apresentaram EMAn inferior ao tratamento controle (sem fitase e 0,41% de Pd) para a fase de crescimento, mesmo com aves alojadas em sala de metabolismo com temperatura controlada (25 ± 3 °C). Os níveis de fitase não influenciaram o coeficiente de metabolizabilidade da matéria seca (P>0,05) de ambas as fases, assemelhando-se aos resultados observados por Fukayama et al. (2008), que não encontraram diferença no CMMS quando avaliaram dietas controle positivo (sem fitase e 0,42% de Pd) e controle negativo (0,27% de Pd) com 500; 750 e 1000 FTU/kg, para o período de 1 a 20 dias.
O consumo e o coeficiente de retenção aparente do nitrogénio (CRN) para a fase inicial (Quadro 3) apresentaram redução linear, de acordo com as equações:
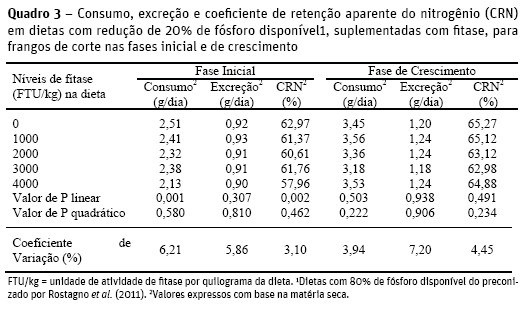
y = 2,506 – 0,0000782x (R2 = 0,786; P<0,01) e y = 62,8632 – 0,0009646x (R2 = 0,664; P<0,01), respectivamente. Os níveis crescentes de fitase nas dietas proporcionaram redução no consumo de nitrogénio, provavelmente devido à redução da formação de complexos fitato-enzimas proteolíticas e fitato-aminoácidos, permitindo maior disponibilização de aminoácidos da proteína intacta e maior aproveitamento dos aminoácidos adicionados à dieta, contribuindo para o atendimento da exigência das aves.
A redução do CRN ocorreu devido à redução do consumo de nitrogénio pelas aves. A excreção desse mineral não foi afetada (P>0,05) pelos níveis de fitase na dieta decido às reduções do consumo e do coeficiente de retenção.
Para a fase de crescimento não houve efeito dos níveis de fitase sobre o consumo, excreção e coeficiente de retenção aparente do nitrogénio (P>0,05) (Quadro 3). Estes resultados assemelham-se aos observados por Gomide et al. (2011) para o período de 28 a 35 dias, em que suplementação de fitase não influenciou o balanço e a retenção de nitrogénio. Para estes autores, a maneira eficiente de reduzir a excreção de nitrogénio e melhorar o CRN, sem prejudicar o desempenho das aves, é por redução da proteína bruta, com inclusão de aminoácidos industriais à dieta.
Não houve efeito dos níveis de fitase (P>0,05) sobre o consumo, excreção e coeficiente de retenção aparente do cálcio (CRCa) para a fase inicial (Quadro 4).
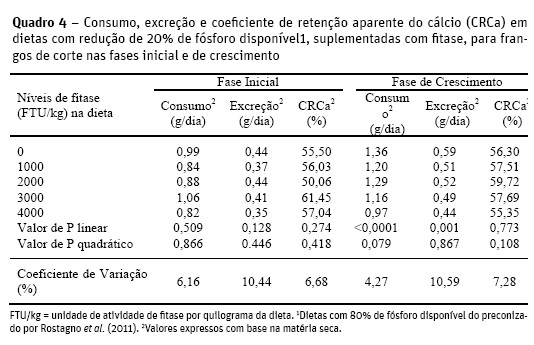
Shirley e Edwards Junior (2003) destacaram que a concentração fisiológica de cálcio em aves, é mais difícil de ser regulada que a de fósforo, e que dietascom mais de 3000 FTU/kg proporcionaram pouco efeito na retenção de Ca.
Silva et al. (2008), verificaram que os níveis de PB (15 e 17%) e Pd (0,25 e 0,34%) em dietas suplementadas com fitase (500 FTU/kg), não reduziram as excreções de Ca, e os CRCa foram iguais ou menores que o da dieta controle, para frangos de corte na fase de 1 a 21 dias.
Por outro lado, Oliveira et al. (2008) avaliaram dietas com dois níveis de fitase (0 e 25 FTU/kg) e três disníveis de Pd (0,31%; 0,38% e 0,45%) em aves do 13º ao 22º dia de vida e constataram aumento no CRCa em todos os níveis de Pd com fitase, em relação à dieta sem fitase.
Estas discrepâncias de resultados demonstram a necessidade do conhecimento da máxima redução possível de Pd e da melhor relação Ca:Pd, em dietas suplementadas com fitase, para que seja alcançado a máxima eficiência da enzima, sem comprometer o desempenho das aves.
Na fase de crescimento a elevação dos níveis de fitase reduziu linearmente o consumo e a excreção de cálcio, respectivamente, segundo as equações: y = 1,36 – 0,0000822x (R2 = 0,763; P<0,01) e y = 0,5772 – 0,0000334x (R2 = 0,867; P<0,01).
A redução no consumo de cálcio, pode estar associada à melhor relação fisiológica cálcio:fósforo para esta fase. A necessidade da adequação no consumo decorreu da ação da fitase em disponibilizar cálcio complexado, e do atendimento das exigências das aves em relação a esse mineral. A redução da excreção de cálcio está relacionada à redução do consumo, e devido as reduções paralelas do consumo e excreção, o CRCa não foi influenciado (P>0,05) pelos níveis de fitase.
Os níveis de inclusão de fitase não influenciaram o consumo de fósforo (P>0,05) das aves na fase inicial (Quadro 5), mas proporcionaram redução linear na excreção: y = 0,1972 – 0,0000092x (R2 = 0,49; P<0,05) e elevação linear no coeficiente de retenção aparente deste mineral: y = 62,5372 + 0,002102x (R2 = 0,662; P<0,01), indicando que a fitase foi eficiente em disponibilizar fósforo do ácido fítico dos ingredientes das dietas, e que as aves utilizaram o fósforo retido em suas exigências.
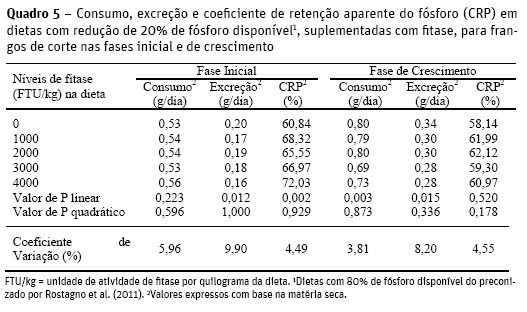
Dietas com baixo nível de Pd, suplementadas com fitase, apresentam maior proporção de fósforo fítico por unidade de fósforo total e elevam a eficiência da fitase em hidrolisar o fitato (Meneghetti et al., 2011). Silva et al. (2008) relataram que a suplementação de fitase na dieta (500 FTU/kg) proporcionou redução de 50% na excreção de fósforo e melhoria de 20% no coeficiente de retenção deste mineral em relação à dieta controle. Similarmente, Lelis et al. (2010) observaram em frangos de corte de 14 a 24 dias de idade, em dieta suplementada com fitase (500 FTU/ kg), redução de 39,97% na excreção de fósforo, e melhoria na retenção em 43,93%, em relação à mesma dieta sem fitase.
Artifícios nutricionais que reduzam a excreção, e melhoram a retenção do fósforo dos ingredientes de origem vegetal das dietas, são benéficos ao ambiente, sobretudo em regiões com intensa atividade avícola.
Para a fase de crescimento, os níveis de inclusão de fitase nas dietas proporcionaram redução linear no consumo: y = 0,808 – 0,0000238 (R2 = 0,583; P<0,01) e na excreção de fósforo: y =0,3252 – 0,0000122x (R2 = 0,816; P<0,05), sem influenciar o CRP (P>0,05), devido as reduções simultâneas no consumo e excreção. A diferença na resposta das aves desta fase em relação à fase inicial pode decorrer da menor exigência em fósforo, e da maior maturidade e eficiência do trato digetório das aves de maior idade em degradar complexos, minimizando os efeitos da fitase.
Gomide et al. (2011) observaram redução no consumo de fósforo em aves de 28 a 35 dias de idade, quando submetidas a dietas com níveis reduzidos de Pd, e afirmaram que a redução de Pd nas dietas suplementadas com fitase é eficiente em reduzir a excreção de fósforo das aves sem afetar o desempenho.
Meneghetti et al. (2011) reportaram que dietas suplementadas com fitase melhoraram o CRP em relação à dieta sem fitase, em frangos de corte na fase inicial. No entanto, não verificaram efeitos dos altos níveis de fitase (1500 a 10000 FTU/kg) sobre esta variável, para a fase de crescimento (22 a 35 dias).
Conclusões
A suplementação de fitase até 4000 FTU/kg em dietas a base de milho e farelo de soja, para frangos de corte em estresse por calor, reduz a energia metabolizável aparente corrigida para o balanço de nitrogénio, diminui a excreção e melhora o coeficiente de retenção do fósforo, na fase inicial. Para a fase de crescimento, há redução na energia metabolizável aparente corrigida para o balanço de nitrogénio e diminuição na excreção de cálcio e fósforo.
Agradecimentos
Os autores agradecem ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pelo financiamento da pesquisa.
Comitê de Ética em Experimentação Animal
O protocolo de ensaio animal está de acordo com CEEA e foi aprovado pelo comitê de ética no uso de animais da Universidade Federal do Piauí pelo parecer nº 044/09.
Referências Bibliográficas
A viagen: Ross broiler management manual (2009). (acesso em 11 de setembro de 2014). Disponível em: http://pt.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross_Broiler_Manual_09.pdf. [ Links ]
Buso, W.H.D.; Morgado, H.S. e Machado, A.S. (2011) - Fitase na alimentação de frangos de corte. PUBVET, vol. 5, n. 36, Ed.183, art.1232. [ Links ]
Cella, P.S.; Donzele, J.L.; Oliveira, R.F.M.; Albino, L.F.T.; Ferreira, A.S.; Gomes, P.C.; Valerio, S.R. e Apolonio, L.R. (2001) - Níveis de lisina mantendo a relação aminoacídica para frangos de corte no período de 1 a 21 dias de idade, em diferentes ambientes térmicos. Revista Brasileira de Zootecnia, vol. 30, n. 2, p. 433-439. [ Links ]
Feitosa, S.M.R. (2010) -Alterações climáticas em Teresina-PI decorrentes da urbanização e supressão de áreas verdes. Dissertação (Mestrado). Curso de Pós-graduação em Desenvolvimento e Meio Ambiente, Universidade Federal do Piauí, 112f. [ Links ]
Fukayama, E.H; Sakomura, N.K.; Dourado, L.R.B.; Neme, R.; Fernandes, J.B.K. e Marcato, S.M. (2008) - Efeito da suplementação de fitase sobre o desempenho e a digestibilidade dos nutrientes em frangos de corte. Revista Brasileira de Zootecnia, vol. 37, n. 4, p. 629-635. [ Links ]
Gomide, E.M.; Rodrigues, P.B.; Bertechini, A.G.; Freitas, R.T.F.; Fassani, E.J.; Reis, M.P.; Rodrgues, N.E.B. e Almeida, E.C. (2011) - Rações com níveis reduzidos de proteína bruta, cálcio e fósforo com fitase e aminoácidos para frangos de corte. Re-vista Brasileira de Zootecnia, vol. 40, n. 11, p. 24052414. [ Links ]
Laurentiz, A.C.; Junqueira, O.M.; Filardi, R.S.; Duarte, K.F.; Assuena, V. e Sgavioli, S. (2009) - Desempenho, composição da cama, das tíbias, do fígado e das excretas de frangos de corte alimentados com rações contendo fitase e baixos níveis de fósforo. Revista Brasileira de Zootecnia, vol. 38, n. 10, p. 1938-1947. [ Links ]
Lelis, G.R.; Albino, L.F.T.; Tavernari, F.C. e Rostagno, H.S. (2009) - Suplementação dietética de fitase em dietas para frangos de corte. Revista Eletrônica Nutritime, vol. 6, n. 2, p. 875-889. [ Links ]
Lelis, G.R.; Albino, L.F.T.; Silva, C.R.; Rostagno, H.S.; Gomes, P.C. e BORSATTO, C.G. (2010) - Suplementação dietética de fitase sobre o metabolismo de nutrientes de frangos de corte. Revista Brasileira de Zootecnia, vol. 39, n. 8, p. 1768-1773. [ Links ]
Meneghetti, C.; Bertechini, A.G.; Rodrigues, P.B.; Fassani, E.J.; Brito, J.A.G.; Reis, M.P. e Garcia, Jr, A.A.P. (2011) - Altos níveis de fitase em rações para frangos de corte. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, vol. 63, n. 3, p. 624-632. [ Links ]
Mitchel, R.D. e Edwards Jr, H.M. (1996) -Effects of phytase and 1,25-didroxicholecalciferol on phytase and utilization and quantitative requirement for calcium and phosphorus in young broiler chickens. Poultry Science, vol. 75, p. 111-119. [ Links ]
Oliveira, G.A.; Oliveira, R.F.M.; Donzele, J.L.; Cecon, P.R.; Vaz, R.G.M.V. e Orlando, U.A.D. (2006a) - Efeito da temperatura ambiente sobre o o desempenho e as características de carcaça de frangos de corte dos 22 a 42 dias. Revista Brasileira de Zootecnia, vol. 35, n. 4, p. 1398-1405. [ Links ]
Oliveira, R.F.M.; Donzele, J.L.; Abreu, M.L.T.; Ferreira, R.A.; Vaz, R.G.M.V. e Cella, P.S. (2006b) - Efeitos da temperatura e da umidade relativa sobre o desempenho e o rendimento de cortes nobres de frangos de corte de 1 a 49 dias de idade. Revista Brasileira de Zootecnia, vol. 35, n. 3, p. 797-803.
Oliveira, M.C; Gravena, R.A. Marques, R.H.; Guandolini, G.C. e Moraes, V.M.B. (2008) -Utilização de nutrientes em frangos alimentados com fitase e níveis reduzidos de fósforo náo-fítico. Arquivo Brasileiro de Medicina Veterinária e Zootecnica, vol. 60, n. 2, p. 436-441. [ Links ]
Rostagno, H.S.; Albino, L.F.T.; Donzele, J.L.; Gomes, C.G.; Oliveira, R.F.; Lopes, D.C.; Ferreira, A.S.; Barreto, S.L.T. e Euclides, R.F. (2011) -Quadros brasileiras para aves e suínos: composição de alimentos e exigências nutricionais. Viçosa: UFV, Departamento de Zootecnia, 3 ed. 252p. [ Links ]
Santos, L.M.; Rodrigues, P.B.; Alvarenga, R.R.; Naves, L.P.; Hespanhol, R.; Lima, G.F.R.; Lara, M.C.C. e Silva, L.R. (2011) -Níveis de fósforo disponível e cálcio em rações suplementadas com fitase para frangos de corte nas fases de crescimento e final. Revista Brasileira de Zootecnia, vol. 40, n. 11, p. 2486-2495. [ Links ]
Selle, P.H. e Ravindran, G. (2007) - Microbial phytase in poultry nutrition. Animal Feed Science and Technology, vol. 135, p.1-4. [ Links ]
Shirley, R.B. e Edwards Junior, J.A. (2003) - Graded levels of phytase past industry standards improves broilers performances. Poultry Science, vol. 82, p. 671-680. [ Links ]
Shoener, F.J.; Hoppe, P.P.; Scwarz, G.; Wiesche, H. (1993) -Comparison of microbial phytase and inorganic phosphate in male chickens – the influence on performance data, mineral retention and dietary calcium. Journal of Animal Physiology Nutrition, vol. 69, n. 5, p. 235-244. [ Links ]
Silva, D.J. e Queiroz, A.C. (2002) - Análise de alimentos: métodos químicos e biológicos. 3 ed., Viçosa: UFV, 235p. [ Links ]
Silva, Y.L.; Rodrigues, P.B.; Freitas, R.T.F.; Bertechini, A.G.; Fialho, E.T.; Fassani, E.J. e Pereira, C.R. (2006) - Redução de proteína e fósforo em rações com fitase para frangos de corte no período de 1 a 21 dias de idade. Desempenho e teores de minerais na cama. Revista Brasileira de Zootecnia, vol. 35, n. 3, p. 840-848. [ Links ]
Silva, Y.L.; Rodrigues, P.B.; Freitas, R.T.F.; Zangeronimo, M.G. e Fialho, E.T. (2008) - Níveis de proteína e fósforo em rações com fitase para frangos de corte, na fase de 14 a 21 dias de idade. 2. Valores energéticos e digestibilidade de nutrientes. Revista Brasileira de Zootecnia, vol. 37, n. 3, p. 469477. [ Links ]
Silva, Y.L.; Rodrigues, P.B.; Zangeronimo, M.G.; Fialho, E.T.; Freitas, R.T.F. e Alvarenga, R.R. (2012) -Redução de proteína e fósforo em dietas com fitase para frangos de corte dos 22 a 42 dias de idade. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, vol. 64, n. 1, p. 127-136. [ Links ]
Statistical Analysis System – SAS. (1996) -SAS/ STAT: users guide, version 6.11 ed. Cary, 842p. [ Links ]
Teixeira, E.N.M; Silva, J.H.V.; Goulart, C.C.; Filho, J.J. e Ribeiro, M.L.G. (2013) - Suplementação da fitase em rações com diferentes níveis de fósforo disponível para frangos de corte. Revista Ciência Agronômica, vol. 44, n. 2, p. 390-397. [ Links ]
Welker, J.S.; Rosa, A.P.; Moura, D.J.; Machado, L.P.; Catelan, F. e Uttpatel, R. (2008) -Temperatura corporal de frangos de cortes em diferentes sistemas de climatização. Revista Brasileira de Zootecnia, vol. 37, n. 8, p. 1463-1467. [ Links ]
Recebido/Received: 2014.03.11
Aceite/Accepted: 2014.08.18

