










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.37 no.4 Lisboa dez. 2014
REVISÃO
Leptina e grelina na produção de ruminantes
Leptin and ghrelin in ruminant production
Luciana H. Kowalski1, José A. Freitas2, Sergio R. Fernandes3, Paulo R. Junior4, Jovanir I. M. Fernandes5 e Marina G. Berchiol da Silva6
1 Universidade Federal do Paraná - Campus Palotina; Rua Pioneiro, 1509, Palotina - PR, Brasil; 85950-000. E-mail: lucianahelenak@gmail.com, author for correspondence
2 Universidade Federal do Paraná - Campus Palotina; Rua Pioneiro, 2153, Jardim Dallas, Palotina - PR, Brasil; 85950-000. E-mail: freitasjaf@ufpr.br
3 Universidade Federal do Paraná- Campus Curitiba-PR. Rua Professor Ewaldo Schiebler, 1200, Jardim Social, Curitiba-PR, Brasil; 82530-160. E-mail: srfernandes83@gmail.com
4 Universidade Federal do Paraná- Campus Curitiba-PR. Rua dos Funcionários, 1540, Juvevê, Curitiba-PR, Brasil; 80035-050. E-mail: parossi@ufpr.br
5 Universidade Federal do Paraná- Campus Palotina. Rua Pioneiro, 2153, Jardim Dallas, Palotina- PR, Brasil; 85050-000. E-mail: jovanirfernandes@gmail.com
6 Faculdade Eduvale/Avaré-SP., Av. Prefeito Misael Eufrásio Leal, 347, Jardim América, Avaré-SP; 18705-050. E-mail: gabiberchiol@hotmail.com
RESUMO
A leptina e a grelina são hormonas peptídicas que foram estudados primeiramente em humanos, devido aos problemas de obesidade e diabetes. Estas hormonas estão presentes em vários mamíferos, incluindo os animais domésticos. As funções conhecidas da leptina em ruminantes estão relacionadas com a regulação do consumo alimentar, a sinalização da condição corporal para a função reprodutiva e ao estímulo da resposta imune. Os polimorfismos do gene da leptina favorecem a seleção de animais com melhores características produtivas, pois estão associados a características qualitativas do leite (teor de gordura e proteína) e da carcaça (espessura de gordura subcutânea e marmoreio). A grelina foi bastante estudada em humanos, havendo poucas funções da grelina descritas em animais de produção. Porém, a grelina atua juntamente com a leptina na regulação do consumo alimentar, e existem evidências de que as duas hormonas regulam, também, a reprodução. Nesse contexto, o objetivo desta revisão consistiu em relatar as funções conhecidas da leptina e da grelina em ruminantes, e a influência destas hormonas nas características produtivas destes animais.
Palavras-chave: bovinos, características qualitativas, consumo alimentar, reprodução
ABSTRACT
Leptin and ghrelin are protein hormones which were studied first in humans due to diseases as obesity and diabetes.
These hormones are present in various mammals, including domestic animals. The known functions of leptin in the ruminant production are related to feed intake regulation, body status indication to the reproductive function and immune response stimulus. The polymorphisms of leptin gene can be used in the selection of animals with better productive traits, because they are associated to milk qualitative traits (fat and protein content) and carcass (backfat thickness and marble score). Ghrelin was widely studied in humans, and few functions of this hormone were reported for domestic animals. However, ghrelin is associated to leptin and both hormones have important function on feed intake regulation, and some evidences indicate that also affect reproduction. In this context, the objective of this review was to report leptin and ghrelin functions in ruminants and the influence of these hormones on productive traits of these animals.
Keywords: cattle, feed intake, qualitative traits, reproduction
Introdução
Desde a descoberta das hormonas leptina e grelina que houve considerável avanço na pesquisa sobre suas funções em animais e humanos (Kojima et al., 1999; Negrão e Licino, 2000). Porém, as funções mais estudadas e elucidadas estão relacionadas à regulação do consumo alimentar e à diminuição de peso corporal, que foram o foco dos primeiros estudos com leptina e grelina (Zhang et al., 1994; Kojima et al., 1999).
Há grande interesse no estudo da leptina em humanos, pois esta hormona pode ser utilizada no tratamento da obesidade, assim como pode ser responsável pela predisposição à obesidade (Hervey, 1959; Zhang et al., 1994; Campfield et al., 1995). Outra função importante e muito estudada em humanos está relacionada com a ação da leptina na reprodução.
Esta hormona atua como sinalizador da condição corporal e regula a cascata hormonal que dá início à pulsatilidade da hormona libertadora de gonadotrofinas – GnRH (Hausman et al., 2012). Os estudos iniciais com a grelina tiveram foco na função desta hormona no estímulo da hormona do crescimento (GH). Posteriormente, estudos demonstraram que a grelina atua em conjunto com a leptina na regulação do consumo de alimentos. A partir daí, esses duas hormonas tornaram-se foco de estudos sobre controle da obesidade na medicina humana. Nos animais, várias pesquisas têm sido direcionadas para compreender a função da leptina e grelina na regulação do consumo alimentar. Estudos mostraram que esses duas hormonas atuam, também, na reprodução (Sirotkin e Meszarosová, 2009), e que a leptina influencia a produção e a qualidade do leite em bovinos (Chilliard et al., 2001; Sadeghi et al., 2008; Cano et al., 2009), a imunidade (Agarwal et al., 2009) e a qualidade de carcaça (Geary et al., 2003). Perante as descobertas recentes sobre a ação da leptina e da grelina nos animais, principalmente em animais domésticos de produção, o objetivo desta revisão em sintetizar as funções da leptina e da grelina em ruminantes, e a influência destas hormonas nas características produtivas destes animais.
Leptina
Histórico
As primeiras descrições da leptina foram feitas por Hervey (1959) e Coleman (1973), que realizaram experiências de parabiose (união fisiológica de dois organismos) com camundongos. Esses autores relataram a existência de um fator circulante que levava camundongos obesos a emagrecer. No início da década de 1990, Zhang et al. (1994) realizaram a clonagem do "fator circulante" descrito anteriormente, que pela primeira vez ficou conhecido e foi nomeado como leptina, onde leptos deriva do grego e significa magro. Conforme esses autores, a leptina é uma hormona proteico formada por 146 aminoácidos, com massa de 16 quilodáltons (kDa) e que apresenta expressão génica nos adipócitos, sendo sintetizada e armazenada no tecido adiposo. Posteriormente, outros estudos de parabiose foram realizados com camundongos das linhagens ob/ ob e db/db (obesos) e normais (Zhang et al., 1994). Quando a circulação sanguínea dos camundongos ob/ob foi ligada com a dos camundongos normais e db/db, observou-se que os primeiros emagreceram. Além disso, os camundongos normais emagreceram quando tiveram a circulação sanguínea ligada a dos camundongos db/db. Concluiu-se nesses ensaios que os camundongos normais e db/db produziam leptina, enquanto os camundongos ob/ob apresentavam falha na produção deste hormona. Assim, com a circulação interligada, os camundongos ob/ob passavam a ter leptina circulante, o que originava na diminuição do consumo de alimentos e na perda de peso. Já os camundongos db/db apresentavam uma provável falha na expressão do receptor da leptina (Houseknecht et al., 1998). Portanto, apesar de haver leptina circulante nos camundongos db/db, estes permaneceram obesos devido à falha no receptor, na sinalização e interpretação pelo sistema nervoso central (SNC).
Regulação do consumo alimentar e do metabolismo energético
A leptina tem ação periférica e no SNC (Salman e Costa, 2006). Em conjunto, estas duas ações regulam o consumo alimentar, a mobilização de gordura e a captação de glicose (Ceddia et al., 1998; Chilliard et al., 2001).
A leptina é produzida no tecido adiposo. À medida que o peso corporal aumenta ocorre aumento na deposição de tecido adiposo, o que leva ao aumento da quantidade de adipócitos e, portanto, da síntese e liberação de leptina. Essa condição fisiológica afeta o consumo de alimento, que é regulado pela ação da leptina no núcleo arqueado do hipotálamo. O aumento da concentração plasmática de leptina sinaliza a saciedade, levando à diminuição do consumo de alimento. Esta queda no consumo alimentar leva à redução da concentração plasmática de leptina (Schwartz et al., 1996; Friedman e Halaas, 1998; Figura 1).

No núcleo arqueado do hipotálamo existem duas populações de neurónios: os orexígenos, que secretam o neuropeptídeo Y (NPY) e a agouti related protein (AgRP); e os anorexígenos, que secretam o transcrito regulado por cocaína e anfetamina (CART) e o peptídeo de melanocortina (alfa-MSH), que é derivado da propriomelanocortina (POMC). Esses receptores são capazes de regular a ingestão ou privação de alimentos e o gasto energético. O aumento da concentração plasmática de leptina, e a ligação desta hormona aos neurónios orexígenos inibe a atividade dos mesmos, o que diminui a liberação de NPY e AgRP. Nessa condição, a leptina estimula a atividade dos neurónios anorexígenos, o que leva ao aumento da libertação de alfa-MSH e CART, levando à diminuição do consumo alimentar. Quando há diminuição na concentração plasmática de leptina, os neurónios anorexígenos não são estimulados e os neurónios orexígenos deixam de ser inibidos, ocasionando o aumento do consumo alimentar (Ribeiro et al., 2007; Figura 2).
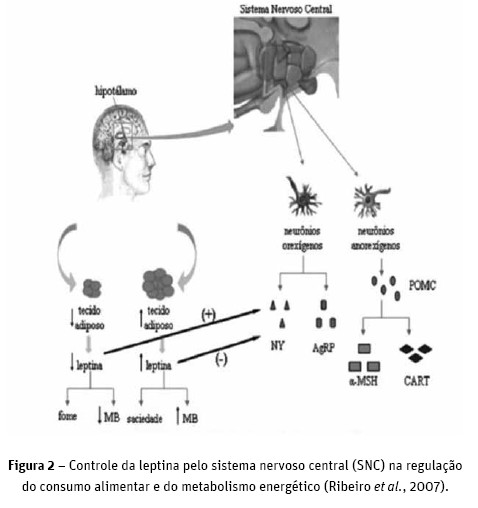
Portanto, a leptina não tem ação direta na regulação do consumo alimentar, mas age por uma cascata de reações hormonais em que o NPY é o principal agente destas reações. Nessa condição, a leptina é considerada um sinalizador da condição corporal.
Reprodução
A maioria dos estudos que relacionam a leptina com a reprodução foi realizada em humanos e camundongos. Sabe-se que a leptina atua na sinalização da condição corporal, o que acarreta no início ou não da puberdade ou do ciclo reprodutivo. As funções desta hormona na reprodução de ruminantes ainda são pouco conhecidas.
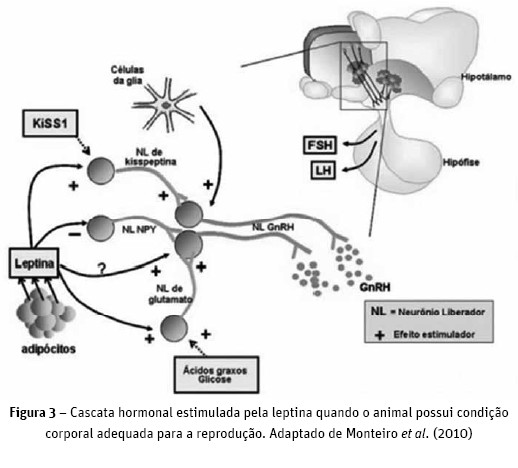
O início da puberdade está intimamente ligado à ingestão de alimento, taxa de crescimento e adiposidade (Williams et al., 2002). Esses três fatores estão relacionados à concentração plasmática de leptina. A principal função da leptina na reprodução é sinalizar a condição corporal (Monteiro et al., 2010). Em animais com condição corporal inadequada, o GnRH apresenta pulsos de baixa frequência e de baixa amplitude, levando à falha no atraso da puberdade em animais jovens e no ciclo reprodutivo de animais adultos. Animais magros apresentam baixa quantidade de gordura corporal, o que leva à baixa síntese e libertação de leptina no plasma. Esta aumenta à medida que ocorre aumento da deposição de tecido adiposo e do tamanho e/ou número de adipócitos.
O NPY é o mediador primário da leptina no hipotálamo, onde altas concentrações de leptina inibem o NPY e estimulam a Kisspeptina (Kiss 1), ocasionando a secreção de GnRH com maior frequência e amplitude (Negrão e Licino, 2000; Quintero e Cortez, 2008; Monteiro et al., 2010; Figura 3).
Imunidade
A função da leptina na imunidade ainda não é total-mente compreendida. Acredita-se que os receptores da leptina sejam da família das citocinas de classe I, que estão relacionadas com o aumento da atividade fagocitária dos macrófagos. Além de regular a atividade dos macrófagos, a leptina induz a proliferação e diferenciação das células do sistema imunitário (Matarese et al., 2005; Agarwal et al., 2009).
Indivíduos com baixa quantidade de tecido adiposo apresentam, consequentemente, baixas concentrações plasmáticas de leptina. Nessa condição, pode haver redução na resposta imune das células Th1, o que aumenta a susceptibilidade a infecções. Indivíduos obesos apresentam elevadas quantidades de tecido adiposo e, assim, altas concentrações plasmáticas de leptina, o que leva à diminuição da responsta à leptina. Nestes indivíduos podemos encontrar resposta imune desregulada, com alteração do balanço de células Th1/Th2, o que pode acarretar uma maior susceptibilidade a infecções e respostas inflamatórias anormais (Matarese et al., 2005).
Qualidade de carcaça
Devido aos mecanismos de ação da leptina e ao seu local de produção, alguns estudos têm procurado correlacionar a concentração plasmática deste hormona com características de qualidade de carcaça. Segundo Geary et al. (2003), a concentração plasmática de leptina tem correlação positiva com escore de marmorização, espessura de gordura subcutânea, grau de deposição de músculo, gordura renal, pélvica e cardíaca, sendo indicada para prever a qualidade de carcaça em bovinos. Porém, num estudo realizado por Gillis et al. (2004) com bovinos de corte mostrou que a concentração plasmática de leptina não tem correlação com as características qualitativas da carcaça como: gordura subcutânea, área de olho de lombo, gordura renal, pélvica ou cardíaca. Nesse estudo, os autores não indicaram a concentração plasmática de leptina como parâmetro para prever a qualidade da carcaça.
Nkrumah et al. (2005) avaliaram dois tipos de polimorfismo do gene da leptina (UASMS2 e UASMS3) em bovinos de corte. Os animais com o polimorfismo foram divididos de acordo com os genótipos que possuíam, sendo eles TT, CT e CC. Observou-se correlação entre os genótipos de leptina e características qualitativas da carcaça. Segundo os autores, animais com genótipo TT dos polimorfismos de leptina apresentam qualidade de carcaça superior aos animais com genótipos CT ou CC. Os animais TT apresentaram valores 39% superiores em concentração plasmática de leptina, 31% a 39% superiores em espessura de gordura e 9% a 13% superiores em escore de marmoreio.
Qualidade do Leite
A melhoria da qualidade do leite mediada pela leptina também depende da seleção de animais com polimorfismos desejáveis. Dentre estes polimorfismos, sabe-se que os genótipos C e T têm relação com a deposição de gordura. Segundo Buchanan et al. (2003), o alelo T tem relação com o aumento da deposição de gordura corporal. Porém, sabe-se que o maior depósito de gordura corporal tem correlação com a quantidade de gordura no leite, principalmente em vacas no início de lactação. Os mesmos resultados foram observados por Sadeghi et al. (2008), onde houve acréscimo de gordura e proteína no leite nos animais com homozigose para o alelo T do gene da leptina (Quadro 1).
Outra função da leptina na melhoria da qualidade do leite é sua ação sobre a contagem de células somáticas (CCS). Segundo Bernotiene et al. (2006), há um aumento da leptina circulante quando o animal se encontra em processos inflamatórios e/ou infecciosos. Por este motivo, Kulig et al. (2010) pesquisaram a influência da leptina na CCS, pois sabe-se que o aumento da CCS indica mastite. Nesta pesquisa os autores relataram que os polimorfismos nos genes R4C CC e Sau3AI TT da leptina levaram à diminuição da CCS em vacas Jersey, o que originou melhoria da qualidade do leite.
Grelina
Estudos envolvendo produção animal e grelina são praticamente inexistentes. Porém nesta revisão vale abordar o tema, pois a grelina tem papel importante na regulação do consumo alimentar, uma vez que age em conjunto com a leptina.
Histórico
Os primeiros estudos com a grelina foram realizados no final da década de 1990 por Kojima et al. (1999), sendo assim, sua descoberta é mais recente que a da leptina. O nome grelina tem origem proto-indo-européia, onde Ghre corresponde no inglês à grow, crescimento. O nome deriva do importante papel que a grelina exerce estimulando a hormona do crescimento (GH), sendo esta a sua primeira função conhecida. A grelina é uma hormona proteica formada por 28 aminoácidos e sintetizada principalmente na mucosa estomacal pelas células GR, podendo ser produzida em menor quantidade no sistema nervoso central, rins, placenta e coração (Kojima et al., 1999; Gualillo et al., 2001). São conhecidas várias funções da grelina em humanos, dentre elas podemos citar i) a regulação de secreções de GH, lactotrófica e corticotrófica; ii) o controle da atividade orexígena (estímulo do consumo de alimento), do gasto energético, da secreção gástrica e da motilidade gástrica; iii) a influência na função endócrina pancreática, no metabolismo da glicose e nas ações cardiovasculares e iv) o efeito antiproliferativo em células neoplásicas (Caminos et al., 2002; Seoane et al., 2004; Vilande Júnior et al., 2012).
Regulação do consumo alimentar
O aumento da concentração plasmática de grelina ocorre principalmente antes da alimentação e/ou em condições de restrição alimentar, o que leva ao estímulo do consumo alimentar. Após a ingestão de alimento, estimulada pelo aumento da concentração plasmática de grelina, ocorre diminuição do gasto energético e aumento das reservas energéticas, sendo assim considerado uma hormona anabólica (Vilande Júnior et al., 2012).
A grelina, assim como a leptina, atua no núcleo arqueado do hipotálamo afim de regular o consumo alimentar. Conforme descrito anteriormente, existem dois grupos de neurónios, os orexígenos (NPY e AgRP) e os anorexígenos (CART e alfa-MSH derivado da POMC). Quando a concentração plasmática de grelina aumenta, os neurónios orexígenos são estimulados, principalmente o NPY, o que leva o animal a alimentar-se. Além disso, o aumento da concentração plasmática da grelina ocasiona a diminuição das concentrações plasmáticas de leptina e insulina. Portanto, a leptina e a grelina são hormonas que têm ações antagónicas na regulação do consumo alimentar (Cummings et al., 2001; Tschöp et al., 2001; Vilande Júnior et al., 2012).
As concentrações plasmáticas de grelina apresentam relação inversa com a massa corporal. Seoane et al. (2004) avaliaram as concentrações plasmáticas de grelina em humanos com índice de massa corporal (IMC) baixo (IMC<18,5), normal (18,5=IMC<25) e elevado (IMC=25). As concentrações de grelina foram elevadas em indivíduos com baixo IMC, apresentando média de 340 pg/mL de grelina e baixa em indivíduos com elevado IMC, apresentando média de 70 pg/mL de grelina. Comprovou-se assim que a grelina pode estimular e/ou inibir o consumo de alimento em função do tamanho corporal do indivíduo.
Secreção de GH
A grelina é um dos principais estimulantes da secreção de GH. Após ser produzida nas células da mucosa estomacal (principal local de síntese), a grelina é libertada na circulação sanguínea e atinge a hipófise. Nesta glândula, a grelina liga-se aos receptores secretagogos GHS-R, estimulando a secreção de GH (Seoane et al., 2004).
A maioria dos estudos que avaliaram a função da grelina na regulação da secreção de GH foram realizados visando a medicina humana. Porém, é provável que esta função seja semelhante em animais e humanos.
Leptina e Grelina
A função de maior interesse na produção animal que envolve a leptina e a grelina é o consumo alimentar. As duas hormonas apresentam funções antagónicas, dado que a grelina estimula os neurónios orexígenos, principalmente o NPY, e a leptina deprime os neurónios orexígenos e estimula os neurónios anorexígenos. Desta forma, a leptina estimula a saciedade e a grelina estimula o consumo de alimento (Figura 4).
A influência que estas hormonas exercem na reprodução também é importante para a produção animal. Sirotkin e Meszarosová (2009) avaliaram a influência da leptina e da grelina nas células da granulosa do ovário de porcas. Houve efeito antagónico destas hormonas sobre a apoptose das células da granulosa, onde a leptina estimulou e a grelina inibiu a apoptose. Já a proliferação de células e a secreção de progesterona foram estimuladas por ambos os hormonas. Concluiu-se que a leptina e a grelina podem atuar em conjunto e apresentar papel importante no desempenho reprodutivo dos animais.
Considerações Finais
Estudos realizados com ruminantes mostram que a leptina atua em funções importantes relacionadas com as características de produção, como a qualidade do leite e da carcaça, estímulo à reprodução e melhoria na resposta imune. Além disso, os polimorfismos do gene da leptina têm potencial para serem utilizados na seleção de animais com características produtivas superiores. Embora tenha sido pouco estudado em animais domésticos, a grelina é uma hormona que atua em conjunto com a leptina na regulação do consumo alimentar e na reprodução. Ainda, a sua função relacionada com o controle da secreção de GH torna a grelina uma hormona de grande importância para a fisiologia animal. Assim, os seus mecanismos de ação relacionados com estas três funções - consumo alimentar, reprodução e secreção de GH - devem ser esclarecidos em ruminantes, uma vez que este conhecimento poderá viabilizar o desenvolvimento de novas alternativas de criação que visem explorar o potencial produtivo destes animais.
Referências bibliográficas
Agarwal, R.; Rout, P.K. e Singh, S.K. (2009) - Leptin: a biomolecule for enhancing livestock productivity. Indian Journal of Biotechnology, vol. 8, n. 2, p.169-176. [ Links ]
Bernotiene, E.; Palmer, G. e Gabay, C. (2006) - The role of leptin in innate and adaptive immune responses. Arthritis Research & Therapy, vol. 8, p. 217-226. [ Links ]
Buchanan, F.C.; Van Kessel, A.G.; Waldner, C.; Christensen, D.A.; Laarveld, B. e Schmutz, S.M. (2003) - Hot topic: An association between a leptin single nucleotide polymorphism and milk and protein yield. Journal of Dairy Science, vol. 86, p. 3164-3166. [ Links ]
Caminos, J.E.; Seoane, L.M.; Tovar, S.A.; Casanueva,F.F. e Dieguez, C. (2002) -Influence of thyroid status and growth hormone deficiency on ghrelin. European Journal of Endocrinology, vol. 147, p. 159–163. [ Links ]
Campfield, L.A.; Smith, F.J.; Guisez, Y.; Devos, R. e Burn, P. (1995) - Recombinant mouse OB protein: evidence for a peripheral signal linking adiposity and central neural networks. Science, vol. 269, p. 546-549. [ Links ]
Cano, M.P.G.; Van Nieuwenhove, C.; Chaila, Z.; Bazán, C. e González, S. (2009) -Effects of short-term mild calorie restriction diet and renutrition with ruminant milks on leptin levels and other metabolic parameters in mice. Nutrition, vol. 25, p. 322-329. [ Links ]
Ceddia, R.B.; William Jr., W.N.; Lima, F.B.; Carpinelli, A.R. e Curi, R. (1998) - Pivotal role of leptin in insulin effects. Brazilian Journal of Medical and Biological Research, vol. 31, p. 715-722. [ Links ]
Chilliard, Y.; Bonnet, M.; Delavaud, C.; Faulconnier, Y.; Leroux, C.; Djiane, J. e Bocquier, F. (2001) - Leptin in ruminants. Gene expression in adipose tissue and mammary gland, and regulation of plasma concentration. Domestic Animal Endocrinology, vol. 21, n. 4, p. 271-295. [ Links ]
Coleman, D.L. (1973) - Effects of parabiosis of obese with diabetes and normal mice. Diabetologia, vol. 9, p. 294-298. [ Links ]
Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E. e Weigle, D.S. (2001) - Pre-prandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes, vol. 50, p. 1714-1719. [ Links ]
Friedman, J.M.; Halaas, J.L. (1998) - Leptin and the regulation of body weight in mammals. Nature, vol. 395, p. 763-770. [ Links ]
Geary, T.W.; McFadin, E.L.; MacNeil, M.D.; Grings, E.E.; Short, R.E.; Funston, R.N. e Keisler, D.H. (2003) - Leptin as a predictor of carcass composition in beef cattle. Journal of Animal Science, vol. 81, p. 1-8. [ Links ]
Gillis, M.H., Duckett, S.K.; Sackmann, J.R.; Realini, C.E.; Keisler, D.H. e Pringle, T.D. (2004) - Effects of supplemental rumen-protected conjugated linoleic acid or linoleic acid on feedlot performance, carcass quality, and leptin concentrations in beef cattle. Journal of Animal Science, vol. 82, p. 851-859. [ Links ]
Gualillo, O.; Caminos, J.; Blanco, M.; Garcìa-Caballero, T.; Kojima, M.; Kangawa, K.; Dieguez, C. e Casanueva, F. (2001) - Ghrelin, a novel placental-derived hormone. Endocrinology, vol. 142, n. 2, p. 788-94. [ Links ]
Houseknecht, K.L., Heuvel, J.P.V., Moya-Camarena, S.Y., Portocarrero, C.P., Peck, L.W., Nickel, K.P. e Belury, M. (1998) Dietary conjugated linoleicacid normalizes impaired glucose tolerance in the Zucker diabetic fa/fa rat. Biochem. Biophys. Res. Commun., vol. 244, p. 678-682. [ Links ]
Hausman, G.J.; Barba, C.R. e Lents, C.A. (2012) -Leptin and reproductive function. Biochimie, vol. 94, p. 2075-2081. [ Links ]
Hervey, G.R. (1959) - The effects of lesions in the hypothalamus in parabiotic rats. Journal Physiology, vol. 145, p. 336-352. [ Links ]
Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H. e Kangawa, K. (1999) - Ghrelin is a GH releasing acylated peptide from stomach. Nature, vol. 402, p.656 – 660. [ Links ]
Kulig, H.; Kmiec, M. e Wojdak-Maksymiec, K. (2010) -Associations between leptin gene polymorphisms and somatic cell count in milk of Jersey cows. Acta Veterinaria BRNO, vol. 79, p. 237-242. [ Links ]
Matarese, G.; Moschos S. e Mantzoros C.S. (2005) – Leptin in immunology. J. Immunol., vol. 174, p. 3137-3142. [ Links ]
Monteiro, C.D.; Bicudo, S.D. e Toma, H.S. (2010) - Puberdade em fêmeas ovinas. PUBVET, vol. 4, n. 21, art. 856. [ Links ]
Negrão, A.B. e Licinio, J. (2000) - Leptina: o diálogo entre adipócitos e neurónios. Arquivo Brasileiro de Endocrinologia e Metabolismo, vol. 44, n. 3, p. 205214. [ Links ]
Nkrumah, J.D.; Li, C.; Yu, J.; Hansen, C.; Keisler, D.H. e Moore, S.S. (2005) -Polymorphisms in the bovine leptin promoter associated with serum leptin concentration, growth, feed intake, feeding behavior, and measures of carcass merit. Journal of Animal Science, vol. 83, p. 20-28. [ Links ]
Quintero, J.C. e Cortez, Z.T.R. (2008) - Efectos de la leptina en el inicio de la pubertad en animales machos. Revista Colombiana de Ciências Pecuárias, vol. 21, p. 97-108. [ Links ]
Ribeiro, S.M.L.; Santos, Z.A.; Silva, R.J.; Louzada, E.; Donato Junior, J. e Tirapegui, J. (2007) - Leptina: Aspectos sobre o balanço energético, exercício físico e amenorréia do esforço. Arquivo Brasileiro de Endocrinologia e Metabolismo, vol. 51, n. 1, p. 11-24. [ Links ]
Sadeghia, M.; Babaka, M.M.S.; Rahimia, G. e Javaremia, A.N. (2008) - Effect of leptin gene polymorphism on the breeding value of milk production traits in Iranian Holstein. Animal, vol. 2, n. 7, p. 999-1002. [ Links ]
Salman, A.K.D. e Costa, R.B. (2006) - Ação hormonal da leptina em ruminantes. Documentos 107. Porto Velho-RO: Embrapa Rondônia. [ Links ]
Schwartz, M.W.; Baskin, D.G.; Bukowski, T.R.; Kuijper, J.L.; Foster, D.; Lasser, G.; Prunkard, D.E.; Porte Júnior, D.; Woods, S.C.; Seeley, R.J. e Weigle, D.S. (1996) -Specificity of leptin action on elevated blood glucose levels and hypothalamic neuropeptide Y gene expression in ob/ob mice. Diabetes, vol. 45, n. 4, p. 531-535. [ Links ]
Seoane, L.M.; Lage, M.; Al-Massadi, O.; Diéguez, C e Casanueva, F.F. (2004) - Papel de la Ghrelina en la fisiopatología del comportamiento alimentario. Revista de Medicina de la Universidad de Navarra, vol. 48, n. 2, p. 11-17. [ Links ]
Sirotkin, A.V. e Meszarosová, M. (2010) - Comparison of effects of leptin and ghrelin on porcine ovarian granulosa cells. Domestic Animal Endocrinology, vol. 39, p. 1–9. [ Links ]
Tschöp, M,; Weyer, C.; Tataranni, P.A.; Devanarayan, V.; Ravussin, E. e Heiman, M.L. (2001) - Circulating ghrelin levels are decreased in human obesity. Diabetes, vol. 50, p. 707-709. [ Links ]
Vilande Júnior, A.V.; Gomes, C.F.; Allgayer, N e Lara, G.M. (2012) - A grelina e sua contribuição para obesidade e diabetes mellitus tipo 2. Revista Conhecimento Online, vol. 2. [ Links ]
Williams, G.L.; Amstalden, M.; Garcia, M.R.; Stanko, R.L.; Nizielski, S.E.; Morrison, C.D. e Keisler, D.H. (2002) - Leptin and its role in the central regulation of reproduction in cattle. Domestic Animal Endocrinology, vol. 23, p. 339-349. [ Links ]
Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; e Friedman, J.M. (1994) - Positional cloning of the mouse obese gene and its human homologue. Nature, vol. 372, p. 425-32. [ Links ]
Recebido/Received: 2013.10.03
Aceitação/Accepted: 2014.10.15

