










Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Ciências Agrárias
Print version ISSN 0871-018X
Rev. de Ciências Agrárias vol.37 no.3 Lisboa Sept. 2014
ARTIGO
Diversidade e potencial de utilização dos rizóbios isolados de nódulos de Gliricidia sepium
Diversity and potential use of rhizobia from nodules of Gliricidia sepium
Ligiane A. Florentino1, Adauton V. Rezende1, Aline C. Mesquita1; André R. S. Lima1, Douglas J. Marques1 e José M. Miranda1
1 Laboratório de Microbiologia Agrícola, Instituto de Ciências Agrárias, Universidade José do Rosário Vellano – UNIFENAS. Rodovia MG 179, Km 0 - Campus Universitário, Alfenas - MG, 37130-000. E-mails: ligiane.florentino@unifenas.br, author for correspondence; adauton.rezende@unifenas.br, alinecmesquita@hotmail.com, limastef@hotmail.com, douglasjmarques81@yahoo.com.br, jose.miranda@unifenas.br
RESUMO
Pesquisas envolvendo a seleção de bactérias fixadoras de N2, os rizóbios, para inoculação em leguminosas forrageiras, além de contribuir para a melhoria da qualidade nutricional, favorecem a gestão e conservação das pastagens. Dessa forma, o objetivo do estudo foi analisar a diversidade morfológica, fenotípica e simbiótica de rizóbios oriundos de 30 nódulos radiculares da leguminosa Gliricidia sepium. Foram obtidos 33 isolados bacterianos em meio 79, dos quais oito foram selecionados para estudos de tolerância, em diferentes condições de acidez e salinidade, e as propriedades simbióticas nas leguminosas G. sepium, Phaseolus vulgaris e Crotalaria spectabilis. Os isolados bacterianos apresentaram características morfológicas altamente similares, mas diferiram em relação às propriedades fenotípicas e simbióticas. Em relação à G. sepium, todos os isolados inoculados incluindo a estirpe inoculante, BR 8802, apresentaram resultados similares em relação à capacidade de fornecer azoto para o desenvolvimento vegetal. Observou-se que alguns dos isolados apresentaram maior habilidade em sobreviver em diferentes valores de pH e salinidade quando comparados à estirpe BR 8802, sugerindo-se que estes sejam selecionados para estudos futuros.
Palavras-chave: Fixação biológica azoto, leguminosa forrageira, manejo e conservação das pastagens, seleção estirpes inoculantes
ABSTRACT
Research involving the selection of N2 fixing rhizobia for inoculation of forage legumes can contribute to improve the nutritional quality, and promote the management and conservation of pastures. Thus, the aim of this study was to analyze the morphological, phenotypic and symbiotic diversity of rhizobia derived from 30 root nodules of legume Gliricidia sepium. Thirty three bacterial isolates were obtained in medium 79, of which eight were selected for tolerance studies in different conditions of acidity and salinity and the symbiotic properties in G. sepium legumes, Phaseolus vulgaris and Crotalaria spectabilis. Bacterial isolates showed similar morphological characteristics, but differed in regard to phenotypic and symbiotic properties. Regarding G. sepium, all isolates inoculated including inoculant strain, BR 8802, showed similar ability to provide nitrogen for plant growth. However, some of the isolates showed greater ability to survive in different pH and salinity compared to BR 8802, suggesting that these should be selected for further studies.
Keywords: Biological nitrogen fixation, forage legume, inoculants strains selection, management and conservation of pastures
Introdução
A leguminosa Gliricidia sepium (Jacq.) Steud. é nativa no México, América Central e Norte da América do Sul. Apresenta crescimento rápido, enraizamento profundo, o que proporciona alta tolerância a déficit hídrico, permanecendo verde a maior parte do ano. Possui ampla distribuição ecológica sendo considerada espécie de múltiplos usos: forragem, reflorestamento, adubação verde, cercas vivas, entre outros.
Devido à suas características bromatológicas, com valores de proteína bruta variando de 20 a 30% (Carvalho Filho et al., 1997), G. sepium tem sido utilizada como forrageira para bovinos, ovinos e caprinos. Tem-se destacado principalmente na alimentação de ovinos, onde tem sido utilizada como suplemento proteico (Combellas et al., 1999), uma vez que o consumo in natura pode ser limitante devido à libertação de compostos voláteis, os quais podem provocar toxicidade aos animais (Simons e Stewart, 1994). Entretanto, há relatos do uso de G. sepium como único alimento para ovelhas e cabras em que não foi diagnosticado toxicidade, verificando-se ganho de peso pelos animais (Carew, 1983). Na utilização como forragem, esta leguminosa pode ser cultivada como cerca viva, desempenhando dessa forma, duas importantes funções: alimentação animal e recuperação do solo.
Ambas as funções estão diretamente relacionadas ao fato de G. sepium ser uma leguminosa que estabelece simbiose com as bactérias fixadoras de N2, conhecidas vulgarmente como rizóbios, que podem fornecer o azoto (N) total ou parcial ao crescimento desta espécie, reduzindo os custos de implantação e proporcionando maior acumulação de N na parte aérea e um aporte de matéria orgânica com menor relação C/N ao solo. No entanto, têm sido poucos os estudos relacionados com a diversidade de rizóbios capazes de estabelecer simbiose com esta leguminosa (Acosta-Durán e Martinez-Romero, 2002; Bala et al., 2003; Thiao et al., 2004). Aliado a isso, torna-se importante ressalvar que esses trabalhos não foram desenvolvidos no Brasil. O estudo da diversidade de rizóbios que estabelecem simbiose com G. sepium constitui uma importante ferramenta de seleção de estirpes mais eficientes em fornecer N e adaptadas a diferentes condições edafoclimáticas, uma vez que esta leguminosa apresenta ampla distribuição ecológica.
De acordo com a instrução normativa n° 13 de 2011 do Ministério da Agricultura, Pecuária e Abastecimento, duas estirpes de rizóbios, BR 8801 e BR 8802, pertencentes ao género Rhizobium são aprovadas como inoculantes. Entretanto, deve-se considerar que o Brasil possui grande extensão territorial, apresentando, portanto, grande variabilidade nas condições edafoclimáticas, que, segundo alguns autores podem influenciar diretamente na eficiência da estirpe inoculante (Cordovilla et al., 1999).
O objetivo do estudo foi analisar a diversidade morfológica, fenotípica e simbiótica de rizóbios oriundos de nódulos radiculares da leguminosa Gliricidia sepium.
Material e Métodos
Colheita, isolamento de nódulos e diversidade de bactérias nodulíferas em Gliricidia sepium
As plantas de G. sepium utilizadas neste estudo foram localizadas no município de Poço Fundo, sul de Minas Gerais (21°4651 de latitude sul e 45°5754 de longitude oeste). O cultivo de G. sepium foi em linha para servir de cerca viva e divisão dos piquetes de aruana (Panicum maximum Jacq. cv. Aruana). Na época da colheita dos nódulos, G. sepium apresentava porte arbustivo e por isso, a colheita dos nódulos radiculares foi realizada com o auxílio de um enxadeco e escavou-se cuidadosamente as raízes secundárias a partir do tronco até as raízes mais finas, onde foi encontrado o maior número de nódulos. Após a colheita, os nódulos foram armazenados em sacos plásticos e depois lavados em água corrente, usando uma peneira. Após esse procedimento, foram transferidos e armazenados individualmente em frascos de vidro contendo sílica gel na parte inferior e um chumaço de algodão na parte superior. No laboratório, conduziu-se a desinfestação e o isolamento dos nódulos seguindo a metodologia de Vincent (1970), utilizando-se álcool etílico 95% por 30 segundos, em seguida, em H2O2 por 3 minutos e depois, lavados seis vezes com água destilada estéril, seguido do maceramento destes em meio de cultura 79 (Fred e Waksman, 1928).
Posteriormente, as colónias obtidas foram purificadas e caracterizadas morfologicamente nesse mesmo meio, de acordo com o tempo de aparecimento da colónia isolada (2-3 dias – crescimento rápido; 4-5 dias – crescimento intermediário; 6-10 dias – crescimento lento), alteração do pH do meio de cultura (reação ácida, neutra ou alcalina), tamanho, forma, elevação, superfície, borda e cor da colónia, quantidade (baixa, média ou alta) e consistência dos exopolissacarídeos e absorção de indicador (azul de bromotimol).
Um dendrograma foi construído a partir dessas 11 variáveis morfológicas, utilizando uma matriz bi-nária, a partir da qual foi realizado o agrupamento com o pacote hclust usando o método average, no R 2.13.2 (R Development Core Team, 2011). Representantes de cada grupo de similaridade foram selecionados para os testes de tolerância aos diferentes valores de pH e concentrações salinas e de autenticação e eficiência simbiótica em diferentes hospedeiros.
Tolerância aos diferentes valores de pH e concentrações salinas
Para verificar a tolerância aos diferentes valores de pH, o meio 79 foi ajustado para os seguintes valores de pH: 4,0; 4,5; 5,0; 5,5; 6,0; 6,5 e 7,0 pela adição de HCl ou NaOH (Wei et al., 2008). A análise de tolerância das bactérias nos diferentes valores de pH foi realizada de acordo com a metodologia utilizada por Hara e Oliveira (2004), baseada na utilização de escalas de tolerância conforme o crescimento bacteriano nas estrias de repicagem. Como tratamento-controle, foi usado o meio de cultura 79 com valor de pH igual a 7,0.
Os tratamentos foram distribuídos inteiramente ao acaso, com três repetições. As placas foram incubadas a 28°C por 7 dias, onde foi avaliado o desenvolvimento das colónias nas quatro zonas de repicagem, onde foram atribuídos valores para o crescimento para cada isolado variando de 1 (sem crescimento visível na placa) a 4 (máximo crescimento em todas as zonas). Com base no crescimento em placas de Petri, os isolados foram classificados como sensíveis (S), medianamente tolerantes (MT) ou tolerantes (T) (Hara e Oliveira, 2004).
O teste de tolerância às diferentes concentrações salinas foi realizado por meio da adição de NaCl, que proporcionaram uma concentração final no meio de cultura de: 34,5, 86, 171, 256, 342 e 427 mM. Como tratamento-controle, foi usado o meio de cultura 79 na sua composição original, que possui 1,71 mM de NaCl e valor de pH igual a 7,0. O delineamento utilizado e a forma de avaliação foi como descrito anteriormente para o teste de tolerância aos diferentes valores de pH.
Autenticação e eficiência simbiótica dos isolados bacterianos quanto à capacidade de nodular de diferentes hospedeiros
Os isolados selecionados de acordo com as características morfológicas foram inoculados em G. sepium (hospedeiro de origem), Phaseolus vulgaris (hospedeiro promíscuo) e Crotalaria spectabilis (leguminosa forrageira e também utilizada como adubo verde), como o objetivo de verificar a autenticidade deste isolado em nodular e fornecer N para o desenvolvimento destas leguminosas. As plantas foram cultivadas em frascos de vidro escuro reciclado com capacidade de 600 mL contendo 500 mL de solução nutritiva de Jensen (1942) a ¼ de força. Os frascos de vidro foram preparados conforme Florentino e Moreira (2009) e durante a condução do ensaio não houve reposição de solução.
Antes da plantação as sementes foram desinfestadas superficialmente por imersão em álcool etílico 100% por 30 segundos e posteriormente, em hipoclorito a 2% por 2 minutos, seguido de lavagens sucessivas em água esterilizada. As sementes foram colocadas em placas estéreis contendo algodão humedecido e papel de filtro até o início da germinação para posterior retirada dos tegumentos e plantação nos frascos de vidro.
Os tratamentos constaram da inoculação de oito isolados bacterianos de nódulos de G. sepium (UNIFENAS 02-10; 03-13; 03-14; 03-12; 03-10; 03-22; 03-15 e 03-16) e as estirpes aprovadas como inoculantes pelo MAPA para cada espécie: G. sepium, BR 8802; P. vulgaris, CIAT 899T (ou BR 322) e C. spectabilis, BR 2003. No momento do plantação, cada frasco de vidro recebeu uma semente germinada e, em seguida, cada semente foi inoculada com 1 mL das estirpes cultivadas em meio líquido 79 contendo aproximadamente 109 células. Foram utilizados ainda dois tratamentos controle, um sem N mineral e o outro contendo N mineral, onde se adicionou 35 mg L-1 N-NH4NO3.
Os ensaios foram conduzidos durante 40 dias em delineamento inteiramente casualizado (DIC), com três repetições. Após este período, foram avaliados os seguintes parâmetros: matéria seca da parte aérea (MSPA) e da raiz (MSR) e número de nódulos (NN). Os resultados dos ensaios foram submetidos à análise de variância, empregando-se o programa de análise estatística Sisvar, versão 5.3. As médias dos tratamentos foram comparadas pelo teste de Scott-Knott a 5% de probabilidade.
Eficiência simbiótica dos isolados em Gliricidia sepium em vasos com solo
Para os isolados bacterianos que apresentaram comprovada capacidade de nodular G. sepium, citados no ensaio anterior, foi instalado um ensaio em vasos contendo 5 dm3 de solo para verificar a eficiência simbiótica.
O solo utilizado apresentou as seguintes características: pH (H2O) = 5,6; Al trocável = (cmolc dm-3) = 0,1; Ca (cmolc dm-3) = 0,3; Mg (cmolc dm-3) = 0,2; P (mg dm-3) = 1; K (mg dm-3) = 12; H + Al (cmolc dm-3) = 2,5; Soma de bases (cmolc dm-3) = 0,5; CTC efetiva (cmolc dm-3) = 0,6; CTC a pH 7,0 (cmolc dm-3) = 3; V (%) = 17; m (%) = 16; Matéria orgânica (dag Kg-1) = 0,6; P-rem (mg L-1) = 4. A calagem foi realizada com calcário dolomítico de acordo com o método da elevação da saturação de bases para 60%. O solo foi humedecido e incubado em sacos plásticos pretos, sendo esta feita para aumentar a velocidade de reação do calcário. Após 30 dias, tempo suficiente para a reação do calcário, os macro e micronutrientes foram adicionados seguindo a recomendação de Novais et al. (1991).
A sementeira foi realizada 10 dias após a adubação e foram semeadas quatro sementes por vaso. Depois de germinadas, foi realizado o desbaste de duas plantas e as outras duas plantas foram mantidas durante 40 dias, quando foram avaliados os seguintes parâmetros: matéria seca da parte aérea (MSPA), número (NN) e matéria seca dos nódulos (MSN). O delineamento experimental utilizado foi o DIC com quatro repetições. Os dados de MSPA, NN e MSN foram submetidos à análise de variância, empregando-se o programa de análise estatística Sisvar, versão 5.3. As médias dos tratamentos foram comparadas pelo teste de Scott-Knott a 5% de probabilidade.
Resultados e Discussão
Diversidade de bactérias nodulantes em Gliricidia sepium
Após o isolamento dos nódulos radiculares de G. sepium, foi obtido um total de 33 isolados bacterianos, os quais foram agrupados de acordo com o dendrograma indicado na Figura 1. Todos os isolados apresentaram crescimento rápido, produção de exopolissacarídeos variando de média a alta e a grande maioria produziu reação ácida no meio de cultura. Portanto, observou-se baixa diversidade morfológica entre os isolados sendo formados oito grupos de similaridade. No entanto, esses resultados corroboram com os encontrados noutros estudos (Acosta-Dúran e Martínez-Romero, 2002; Thiao et al., 2004). Estes últimos autores relatam ainda que G. sepium somente estabelece simbiose efetiva com rizóbios de crescimento rápido.
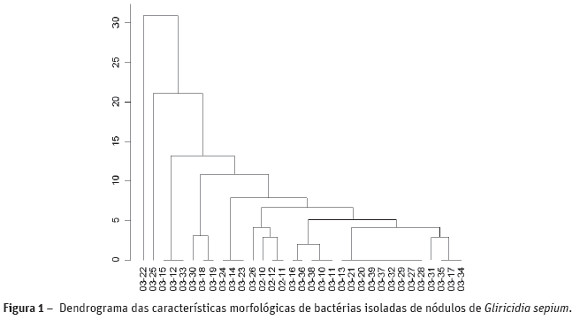
A caracterização morfológica e fenotípica tem sido utilizada para a seleção de estirpes para testes de nodulação, eficiência simbiótica e análises moleculares, uma vez que se torna limitante trabalhar com todos os isolados obtidos dos nódulos das leguminosas em ensaios em estufa (Jaramillo et al., 2013).
Tolerância aos diferentes valores de pH e concentrações salinas
Em relação ao pH, os isolados UNIFENAS 02-10 e 03-16 apresentaram sensibilidade nos valores de pH de 4,5 e 5,0. Já em relação às três estirpes inoculantes, CIAT 899T, BR 8802 e BR 2003, somente a última teve o crescimento reduzido quando cultivada nos valores de pH de 4,5 e 5,0 (Quadro 1).
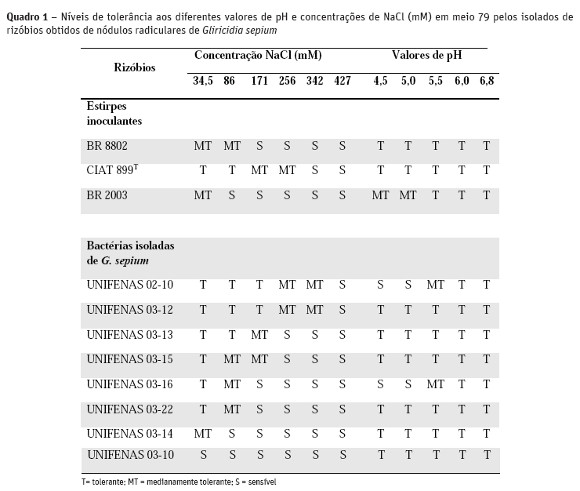
Segundo alguns autores, a maior tolerância à condições de stresse, como acidez e salinidade, está relacionada com alta produção de exopolissacarídeos, os quais atuam como um mecanismo de proteção à célula bacteriana (Bomfeti et al., 2011). No entanto, de acordo com os resultados obtidos, esta generalização nem sempre pode ser verificada.
Em relação a tolerância às diferentes concentrações de NaCl, a máxima concentração tolerada pelas bactérias foi de 342 mM para os isolados UNIFENAS 02-10 e 03-12. No entanto, de acordo com a escala de classificação utilizada, esses isolados foram classificados como moderadamente tolerantes. O máximo crescimento em todas as regiões das placas foi a 171 mM e foi observada para os dois isolados citados anterior-mente. O isolado bacteriano mais sensível foi UNIFENAS 03-10, que não apresentou crescimento na placa na menor concentração testada, 34,5 mM (Quadro 1).
Das estirpes inoculantes, a CIAT 899T foi a mais tolerante, apresentando máximo crescimento até à concentração de 86 mM. As demais, BR 8802 e BR 2003 foram classificadas como medianamente tolerantes na concentração de 34,5 mM.
Relacionando os resultados obtidos no ensaio que avaliou a eficiência simbiótica dos isolados de rizóbios em G. sepium, observa-se que a maior tolerância à salinidade apresentada pelo isolado UNIFENAS 03-13, quando comparada à estirpe inoculante, BR 8802, apresenta relevância para cultivos de G. sepium utilizando a inoculação com estirpes de rizóbios em solos salinos. Estudos comprovam o potencial de utilização desta leguminosa nesses tipos de solos (Farias et al., 2009), os quais podem ser maximizados por meio da inoculação com estirpes de rizóbios eficientes e adaptadas às condições salinas.
Autenticação dos isolados bacterianos quanto à capacidade de nodular diferentes hospedeiros
Em nenhum dos ensaios foi verificado nodulação nos dois controles sem inoculação: com N mineral e sem N mineral, indicando ausência de contaminação. Nos demais tratamentos inoculados com as oito bactérias selecionadas de acordo com as características morfológicas foram observados variabilidade em relação à nodulação e eficiência simbiótica nos diferentes hospedeiros testados: G. sepium, P. vulgaris e C. spectabilis (Quadro 2).
Em G. sepium, cinco dos oito isolados bacterianos estabeleceram simbiose com esta leguminosa, destacando-se UNIFENAS 03-13 e 03-14 que proporcionaram maior desenvolvimento da parte aérea, apresentando resultados superiores à estirpe inoculante BR 8802. Nesse ensaio foi observado ainda que a inoculação com isolados de nódulos de G. sepium, como os dois citados anteriormente e UNIFENAS 03-22, 03-15 e 03-16, sendo este último não nodulífero (Quadro 2), proporcionaram bom desenvolvimento do sistema radicular, o que pode ser atribuído à produção do fitormônio ácido 3-indol acético (AIA) pelo isolado bacteriano, contribuindo para o desenvolvimento do sistema radicular. Em relação a este fato, são encontrados estudos que analisaram a produção de AIA pelas bactérias fixadoras de N2 (Chagas Júnior, 2009). Em P. vulgaris, como em G. sepium, dos oito isolados testados, cinco estabeleceram simbiose com esta leguminosa, sendo que UNIFENAS 02-10 e 03-12 não estabeleceram simbiose com G. sepium (Quadro 2).
O estabelecimento da simbiose está relacionado à libertação de sinais químicos emitidos pelos simbiontes (planta e bactéria), o que explica o fato de ocorrer nodulação numa planta e não noutra (Moreira e Siqueira, 2006). Devido a isso, justifica-se a importância dos estudos de seleção de estirpes para a prática da inoculação no momento da plantação. Dentre os isolados que nodularam P. vulgaris, des-tacam-se UNIFENAS 02-10, 03-13, 03-22 e 03-15, os quais promoveram maior desenvolvimento da parte aérea, superior ao tratamento em que foi utilizada a estirpe inoculante, CIAT 899T, devendo portanto serem incluídas em estudos futuros de eficiência simbiótica com o feijão em vasos e condições de campo. O feijoeiro é uma leguminosa bastante promíscua, ou seja, capaz de estabelecer simbiose com diferentes estirpes de rizóbios, o que dificulta a prática da inoculação no campo, uma vez que o estabelecimento da simbiose irá ocorrer com várias espécies de bactérias nativas (Cardoso et al., 2012). Entretanto, a utilização de estirpes selecionadas tem apresentado resultados satisfatórios em alguns estudos (Ferreira et al., 2009). Já C. spectabilis não apresentou bom desenvolvimento, o que pode ser atribuído à forma de cultivo, uma vez que o tratamento em que foi adicionado N mineral não apresentou desenvolvimento da parte aérea, diferentemente do que foi verificado para as demais leguminosas (Quadro 2).
Eficiência simbiótica dos isolados bacterianos quanto à capacidade de nodular Gliricidia sepium
Neste ensaio foi testada a eficiência simbiótica dos isolados de rizóbios quando inoculados em G. sepium em vasos contendo 5 dm3 solo. Todas as bactérias testadas, incluindo a estirpe inoculante, BR 8802, apresentaram comportamento semelhante (Quadro 3). No entanto, observa-se que a inoculação contribuiu para o maior desenvolvimento da parte aérea quando comparado com o tratamento em que não foi adicionado N mineral, justificando a importância da inoculação.
Também, é importante ressaltar que os resultados obtidos nestes ensaios constituem etapas iniciais para a seleção de isolados eficientes em fixar N2 em simbiose com G. sepium. Portanto, o fato de todos apresentarem a mesma capacidade em fornecer N para o desenvolvimento desta leguminosa nestas condições não deve ser visto como um resultado insatisfatório, uma vez que deverão ser realizados outros estudos com estes isolados, em diferentes condições. Apesar do número reduzido de pesquisas, alguns autores relatam o efeito positivo da inoculação com estirpes de rizóbios em G. sepium (Sanginga et al., 1991).
Estudos preliminares, envolvendo isolamento, caracterização morfológica, fenotípica e testes de nodulação constituem uma importante ferramenta para a seleção de novas estirpes inoculantes. Foi por meio de estudos semelhantes a este que foram identificadas estirpes que atualmente são aprovadas pelo MAPA como inoculantes para culturas de importância agrícola como o feijão-caupi (Lacerda et al., 2004).
Conclusão
Gliricida sepium estabelece simbiose preferencialmente com isolados de rizóbios de crescimento rápido, no entanto, estes apresentam características fenotípicas e simbióticas distintas, as quais apresentam potencial de serem exploradas de acordo com as condições edafoclimáticas predominantes na área de cultivo desta leguminosa.
Agradecimentos
À Fapemig, pelas bolsas de Iniciação Científica aos estudantes do curso de Agronomia, Aline Carvalho de Mesquita e André Ricardo Stefanuto de Lima.
Referências Bibliográficas
Acosta-Durán, C. e Martínez-Romero, E. (2002) - Diversity of rhizobia from nodules of the leguminous tree Gliricidia sepium, a natural host of Rhizobium tropici. Archieves of microbiology, vol. 178, n. 2, p.161-164. [ Links ]
Bala, A.; Murphy, J.P.; Osunde, A.O. e Giller, K.E. (2003) -Nodulation of tree legumes and the ecology of their native rhizobial populations in tropical soils. Applied Soil Ecology, vol. 22, n. 3, p. 211-223. [ Links ]
Bomfeti, C.A.; Florentino, L.A.; Guimarães, A.P.; Cardoso, P.G.; Guerreiro, M.C. e Moreira, F.M.S. (2011) - Exopolysaccharides produced by the symbiotic nitrogen-fixing bacteria of leguminosae. Revista Brasileira de Ciência do Solo, vol. 35, n.3, p. 657-671. [ Links ]
Cardoso, J.D.; Hungria, M. e Andrade, D.S. (2012) -Polyphasic approach for the characterization of rhizobial symbionts effective in fixing N2 with common bean (Phaseolus vulgaris L.). Applied Microbiology and Bbiotechnology, vol. 93, n. 5, p. 2035-2049. [ Links ]
Carew, B.A.R. (1983) - Gliricidia sepium as a sole feed for small ruminants. Tropical Grasslands, vol. 17, n. 4, p. 181-184. [ Links ]
Carvalho Filho, O.M.; Drumond, M.A. e Languidey, P.H. (1997) -Gliricidia sepium. Leguminosa promissora para regiões semi-áridas. Circular Técnica, 35. Petrolina, EMBRAPA-CPATSA. 16 p. [ Links ]
Chagas Júnior, A.F.; Oliveira, L.A. e Oliveira, A.N. (2009) - Produção de ácido indolacético por rizóbios isolados de caupi. Revista Ceres, vol. 56, n. 6, p. 812-817. [ Links ]
Combellas, J.; Rios, L.; Osea, A. e Rojas, J. (1999) -Efecto de la suplementación con follaje de leguminosas sobre la ganancia en peso de corderas recibiendo una dieta basal de pasto de corte. Re-vista de La Facultad de Agronomía, vol. 16, n. 4, p. 211-216. [ Links ]
Cordovilla, D.P.M.; Ligero, F. e Lluch, C. (1999) -Effect of salinity on growth, nodulation and nitrogen assimilation in nodules of faba bean (Vicia faba L.). Applied Soil Ecology, vol. 11, n. 1, p. 1-7. [ Links ]
Farias, S.G.G.; Freire, A.L.O.; Santos, D.R.; Bakke, I.A. e Silva, R.B. (2009) -Efeitos dos estresses hídrico e salino na germinação de sementes de gliricidia [Gliricidia sepium (JACQ.) STEUD.]. Caatinga, vol. 22, n. 4, p. 152-157. [ Links ]
Ferreira, P.A.A.; Silva, M.A.P.; Cassetari, A.; Rufini, M.; Moreira, F.M.S. e Andrade, M.J.B. (2009) -Inoculação com cepas de rizóbio na cultura do feijoeiro. Ciência Rural, vol. 39, n. 7, p. 2210-2212. [ Links ]
Florentino, L.A. e Moreira, F.M.S. (2009) -Características simbióticas e fenotipias de Azorhizobium doebereinerae, microssimbiote de Sesbania virgata. Revista Árvore, vol. 33, n. 2, p. 215-226. [ Links ]
Fred, E.B. e Waksman, S.A. (1928) -Laboratory manual of general microbiology: with special reference to the microorganisms of the soil. New York, McGraw-Hill Book. 145 p. [ Links ]
Hara, F.A.S. e Oliveira, L.A. (2004) - Características fisiológicas e ecológicas de isolados de rizóbios oriundos de solos ácidos e álicos de Presidente Figueiredo, Amazonas. Acta Amazônica, vol. 34, n. 2, p. 343-357. [ Links ]
Jaramillo, P.M.D.; Guimaraes, A.A.; Florentino, L.A.; Silva, K.B.; Nobrega, R.S.A. e Moreira, F.M.S. (2013) -Symbiotic nitrogen-fixing bacterial populationstrapped from soils underagroforestry systems in the Western Amazon. Scientia Agricola, vol. 70, n. 6, p. 397-404. [ Links ]
Jensen, H.L. (1942) -Nitrogen fixation in legouminous plants. I. General characters of root-nodule bacteria isolated from species of Medicago and Trifolium in Australia. Proceedings of the Linnean Society of New South Wales, vol. 66, p. 98-108. [ Links ]
Lacerda, A.M.; Moreira, F.M.S.; Andrade, M.J.B. e Soares, A.L.L. (2004) - Efeito de estirpes de rizóbio sobre a nodulação e produtividade do feijão-caupi. Revista Ceres, vol. 51, n. 293, p. 67-82. [ Links ]
Moreira, F.M.S. e Siqueira, J.O. (2006) - Microbiologia e bioquímica do solo. 2. ed. Lavras, UFLA, 729 p. [ Links ]
Novais, R.F.; Neves, J.C.L. e Barros, N.F. (1991) - Ensaio em ambiente controlado. In: Oliveira, A.J.; Garrido, W.E.; Araújo, J.D. e Lourenço, S. (Eds.) -Métodos de pesquisa em fertilidade do solo. Brasília, EMBRAPA-SAE, p. 189-254. [ Links ]
Sanginga, N.; Manrique, K. e Hardson, G. (1991) -Variation in nodulation and N2 fixation by the Gliricidia sepium/Rhizobium spp. symbiosis in a calcareous soil. Biology and Fertility of Soils, vol. 11, n. 4, p. 273-278. [ Links ]
Simons, A.J. e Stewart, J.L. (1994) - Gliricidia sepium a multipurpose forage tree legume. In: Gutteridge, R.C.; e Shelton, H.M. (Eds.). Forage tree legumes in tropical agriculture. Wallinford, CAB International, p. 30-48. [ Links ]
Thiao, M.; Neyra, M.; Isidore, E. e Sylla, S. (2004) - Diversity and effectiveness of rhizobial strains from Gliricidia sepium native to Reunion Island, Kenya and New Caledonia. World Journal of Microbiology and Biotechnology, vol. 20, n. 7, p. 703-709 [ Links ]
Vincent, J.M.A. (1970) - Manual for the pratical study of root-nodule bacteria. Oxford, Blackwell Scientific Publications. 164 p. [ Links ]
Wei, G.H.; Zhang, Z.X.; Chen, C.; Chen, W.M. e Ju, W.T. (2008) - Phenotypic and genetic diversity of rhizobia isolated from nodules of the legume genera Astragalus, Lespedeza and Hedysarum in northwestern China. Microbiological Research, vol. 163, n. 6, p. 651-662. [ Links ]
Recebido/Received: 2014.05.14
Aceitação/Accepted: 2014.05.21

