










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.36 no.3 Lisboa jul. 2013
Estoque de carbono e atividade microbiana em sistema de plantio direto consolidado no Sul do Brasil
Carbon stocks and microbial activity in a consolidated no tillage system in Southern Brazil
Michael Mazurana1, Jessé Rodrigo Fink1, Estefânia Camargo1, Catiline Schmitt1, Robson Andreazza2 e Flávio Anastácio de Oliveira Camargo1
1 Departamento de Solos, Universidade Federal do Rio Grande do Sul, Bento Gonçalves 7712, Agronomia, 91540-000, Porto Alegre, RS, Brasil.
2 Centro de Engenharias, Universidade Federal de Pelotas, Rua Almirante Barroso 1734, 96010-280, Pelotas, RS, Brasil.E-mail: robsonandreazza@yahoo.com.br, author for correspondence.
RESUMO
A análise da qualidade do solo é uma ferramenta importante para avaliar o ecossistema, bem como planejar a implantação de práticas sustentáveis de manejo. Com isto, o objetivo deste trabalho foi avaliar o estoque de carbono e a atividade microbiana do solo sob diferentes manejos de um experimento de longa duração por meio de amostras de solo deformadas e indeformadas. O experimento está instalado em um Argissolo Vermelho, em sistema de plantio direto há 11 anos com aportes de resíduos vegetais no período de inverno (0, 2, 4, 6 e 8 Mg ha-1) e rotação de culturas no verão (milho/soja ou feijão). No solo foi realizada a determinação do estoque de carbono orgânico do solo (COS) e calculado os coeficientes de humificação (k1) e decomposição (k2) da matéria orgânica do solo. Também foi quantificada a respiração microbiana basal (C-CO2) nas amostras de solo deformadas e indeformadas, os teores de carbono da biomassa microbiana do solo (BMS-C) e o quociente metabólico (qCO ). O COS variou de 21,3 a 27,3 Mg ha-1, sendo que, quantidades de resíduos inferiores a 6 Mg ha-1 adicionados no inverno, mantiveram o COS abaixo da condição original (24,4 Mg ha-1). Os valores obtidos de k1 e k2 foram mais elevados em relação aos demonstrados na literatura para o mesmo tipo de solo. O BMS-C apresentou maior valor para o aporte de 8 Mg ha-1 de resíduos, enquanto que o qCO , embora não significativo, apresenta maior valores para o menor aporte de resíduo. O C-CO2 não apresentou diferença significativa entre amostras deformadas e indeformadas dentro de cada tratamento. No entanto, diferenças significativas entre tratamentos foram observadas nas amostras indeformadas. Embora nossos resultados de atividade microbiana mostrem que a utilização de amostras indeformadas possam ser utilizados para esta determinação, e fornecer melhores resultados, é necessário maiores estudos para o aperfeiçoamento da técnica.
Palavras-Chave: Biomassa microbiana, manejo do solo, qualidade do solo
ABSTRACT
The analysis of soil quality is an important tool to evaluate ecosystems, and to plan and adopt practices of sustainable agricultural management. Thus, the objective of this study was to evaluate the carbon stocks and microbial activity under different management practices in a long-term experiment with disturbed and undisturbed soil samples. The experiment is installed in a Ultisol, in a no tillage system for 11 years with variable inputs of crop residues during winter (0, 2, 4, 6 and 8 Mg ha-1) and in the summer with crop rotation (corn/soybeans or beans). The soil organic carbon stocks (SOC), and the coefficients of humification (k1) and decomposition (k2) of the soil organic matter were determined. The microbial basal respiration (CO2-C) in disturbed and undisturbed soil samples, and the levels of microbial biomass carbon (MBC) and metabolic quotient (qCO2) were also quantified. The SOC ranged from 21.3 to 27.3 Mg ha-1 , whereas the addition of less than 6 Mg ha-1 of crop above ground maintained the SOC below the original condition (24.4 Mg ha-1). The k1 and k2 values were higher than those found in the literature for the same soil. The MBC exhibited higher value to inputs of 8 Mg ha-1 of residues, whereas the qCO , although it was significant higher value for the lowest supply of residue. The CO2-C was not significantly different between disturbed and undisturbed samples in each treatment. However, significant differences between treatments were showed in the undisturbed samples. While our results of microbial activity showed that the use of undisturbed samples can be used for this determination, more studies are needed to improve this technique.
Key-words: Microbial biomass, soil management, soil quality
Introdução
Devido ao aumento dos fluxos de matéria e energia no sistema conservacionista do solo com o sistema de plantio direto há alteração da dinâmica das populações microbianas no solo (Moreira e Siqueira, 2006), dos nutrientes, o fluxo de água e gases, o desenvolvimento do sistema radicular e consequentemente o rendimento das culturas (Santos et al., 2003). Esse aumento permite melhorias nas condições físico-químico-biológicas promovendo maior qualidade ao solo. Segundo Nilsen e Winding (2002), um solo de qualidade pode ser considerado aquele que possui contínua capacidade de funcionar dentro do ecossistema, sustentando a produtividade biológica, mantendo a capacidade ambiental e promovendo o crescimento das plantas. Diversas linhas de pesquisa avaliam a qualidade do solo sob diferentes enfoques como apontado por Sequinatto (2010). Dentre elas, Shukla et al. (2006) apontam a matéria orgânica do solo (MOS) como o melhor indicador da sua qualidade. No entanto, as quantidades de resíduos vegetais mantidos sobre o solo e o tipo de preparo determinam os coeficientes de humificação do material vegetal depositado sobre o solo, bem como, os coeficientes de decomposição da MOS. Quando as taxas de adição de resíduos forem maiores que as taxas de decomposição, há um aumento nos estoques de MOS, favorecendo os processos de agregação do solo levando a estabilidade do sistema.
Os microrganismos do solo também são afetados pela disponibilidade de material orgânico (Moreira e Siqueira, 2006) utilizando os resíduos orgânicos como fonte de carbono, gerando energia para seu desenvolvimento. A quantidade de resíduo vegetal mantido sobre o solo pode levar ao aumento do teor de matéria orgânica, dependendo das condições pedoambientais que afetam os coeficientes de humificação e decomposição (Bayer et al., 2006a), bem como, pode refletir na quantidade e na diversidade da biomassa microbiana no solo.
Por serem sensíveis às mudanças que ocorrem no solo, os microrganismos são importantes indicadores na avaliação das alterações resultantes de diferentes práticas e sistemas de manejo (Pereira et al., 2007; Hungria et al., 2009). Assim, a avaliação da biomassa e respiração microbiana apresenta-se como um importante subsídio para melhor entender a dinâmica dos processos de transformação dos resíduos orgânicos no solo (Petry et al., 2012; Segatto et al., 2012). Entretanto, a interpretação destas avaliações isoladamente nem sempre é satisfatória como um indicador de melhoria na qualidade do solo. Diante disso, a interpretação conjunta de indicadores tem ampliado o entendimento sobre os processos que ocorrem no sistema solo. Neste sentido, a taxa de respiração por unidade de biomassa microbiana (coeficiente metabólico – qCO2) tem sido usada como indicador de eficiência de uso de carbono pela biomassa, ou seja, quanto maior o seu valor, maior a eficiência de acúmulo de carbono pela biota (Balota et al, 1998).
Diversos estudos (Chaer e Tótola, 2007, Trannin et al., 2007) buscam identificar características e comportamento das populações microbianas sob distintas condições de manejo de solo a fim de definir quais condições são mais adequadas para cada ambiente. No entanto, ainda são poucos os estudos que investigam as relações entre microrganismos e o ambiente proporcionado pelo plantio direto. Diante disso, o sistema de plantio direto, por manter resíduos vegetais na superfície do solo, levaria a estimular as populações microbianas devido à maior disponibilidade de material orgânico ao sistema. Além disso, amostras de solo indeformadas, comparadas às deformadas, seriam mais fidedignas às variações de respiração microbiana no sistema de plantio direto, por manterem as características físicas e biológicas das amostras próximas às condições de campo. Em virtude disso, este trabalho objetivou mensurar a respiração microbiana basal (em amostras deformadas e indeformadas), o estoque de carbono orgânico do solo e o carbono da biomassa microbiana, em um experimento de longa duração, sob plantio direto consolidado com distintos aportes de resíduos vegetais no período de inverno e com sucessão de culturas no verão, para caracterizar estes como alguns indicadores da qualidade biológica do solo.
Material e métodos
Experimento de longa duração
O trabalho foi realizado em uma área que está sob sistema de plantio direto desde o ano 2000, cujo solo é classificado como Argissolo vermelho distrófico típico (EMBRAPA, 2006) localizado na Estação Experimental Agronômica da Universidade Federal do Rio Grande do Sul, no município de Eldorado do Sul (coordenadas 30º 05 38.84 S e 51º40 32.35 W). A área experimental é composta por cinco tratamentos, em triplicatas, que se referem a adição de resíduos vegetais da parte aérea de 0, 2, 4, 6 e 8 Mg ha-1 (acrescidos de 30% referente a contribuição do carbono aportado pelas raízes – Zanatta et al., 2007), provenientes da cultura de inverno (Quadro 1).. Durante o verão, foram conduzidos cultivos de grãos apresentados no Quadro 1. A quantidade de resíduos adicionados por estes cultivos foi calculada com base no índice de colheita, sendo estes somados aos resíduos das plantas cultivadas durante o inverno.

Estoque de C, e coeficientes de humificação e decomposição da matéria orgânica
O estoque inicial e atual de carbono no solo foi calculado com base no teor de carbono orgânico total e na densidade do solo, para cada tratamento e mesma camada avaliada no ano de 2000 (0-0,10 m). Os coeficientes de humificação e decomposição da matéria orgânica do solo foram estimados para cada tratamento pelo modelo unicompartimental proposto por Dalal e Mayer (1986), trabalhado por Vieira (2007) e Bayer et al. (2006b). A fixação e emissão de carbono do sistema foi calculado com base nas equações 1 e 2:
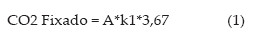
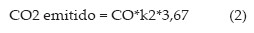
onde, A é a adição anual de carbono (Mg ha-1 ano-1), CO é o carbono orgânico do solo (Mg ha-1), k e k2 são os coeficientes de humificação e decomposição da matéria orgânica, respectivamente e, 3,67 é o fator de conversão de carbono (C) para CO2 (massa molar do C/massa molar de CO2).
Respiração microbiana basal
Para a determinação de respiração microbiana basal, foram escolhidos os tratamentos zero, 4 e 8 Mg ha-1. Essa escolha se deu com base na quantidade média de palha deixada pela cultura da aveia em áreas de lavoura, ou seja, aproximadamente 4 Mg ha-1. A partir de canos de PVC com 7,5 cm de diâmetro e 5,0 cm de altura (profundidade) foram coletadas amostras de solo em entrelinhas de semeaduras, em quadriplicata nas três repetições de cada tratamento, totalizando 36 amostras. Deste total, 18 amostras foram mantidas indeformadas (em tubos de PVC) e, nas restantes, procedeu-se o destorroamento antes de incubá-las, seguindo metodologia proposta por Jenkinson e Powlson (1976). Nas amostras indeformadas isolou-se a parte inferior do tubo com filme plástico de PVC a fim de evitar evolução de CO2 pela parte inferior do cilindro, superestimando a emissão. As amostras de solo foram umedecidas até atingir 60% da capacidade de campo e mantidas em frascos hermeticamente fechados por um período de 86 dias, durante o qual foi determinada a quantidade de CO2 evoluído pela atividade microbiana segundo Jenkinson e Powlson (1976). O período de avaliação se deu com base na taxa de CO2 evoluído pela atividade microbiana, a qual apresentou estabilização a partir de 60 dias após a incubação.
Biomassa microbiana e quociente metabólico(qCO2)
Parte das amostras deformadas, duas repetições de cada tratamento, sofreram fumigação com clorofórmio permanecendo incubadas por 24 horas à temperatura de 26 ± 1 ºC. Duas outras repetições sem fumigação foram adicionadas para a extração do carbono da biomassa microbiana (CBM). A extração do carbono das amostras fumigadas e não fumigadas foi realizada com sulfato de potássio a 0,5 M L-1. Da alíquota filtrada foram utilizados 8 mL para a quantificação do CBM por meio de combustão úmida com oxidação em dicromato de potássio e determinação por titulometria com solução de sulfato ferroso amoniacal. A diferença entre o teor de carbono do solo sem fumigação e do solo fumigado com clorofórmio, determina o carbono da biomassa microbiana. O qCO2 foi calculado pela razão entre a taxa de respiração microbiana basal (RMB) e o carbono da biomassa microbiana (BMS-C) conforme proposto por Anderson e Domsch (1993).
Análise estatística
Os dados foram submetidos à análise estatística pelo software Statiscal Analysis System versão 9.0 (SAS for Windows), e havendo significância da análise de variância compararam-se as médias pelo teste de Tukey (p < 0,05). Também foram realizadas análise de regressão para os dados.
Resultados e discussões
Estoque de carbono orgânico
A adição de resíduos, o estoque de carbono orgânico do solo (COS) e os coeficientes de humificação e decomposição da matéria orgânica (k1 e k2, respectivamente) são apresentados no Quadro 2. A adição anual total de carbono orgânico no solo providos pelas culturas variou de 21,3 a 27,3 Mg ha-1, havendo um aumento no conteúdo de COS com o aumento na quantidade de resíduos adicionados. No entanto, observou-se que adições de resíduos menores que 6Mg ha-1 não conseguiram manter o COS no mesmo nível da condição original (C0), apresentando redução de 12,7, 12,6 e 10,2% para os tratamentos zero, 2, e 4 Mg ha-1 de palha, respectivamente. Isso indica que, para as condições edafoclimáticas do local do estudo, nem sempre há um incremento de carbono ao solo suficiente para manter ou melhorar a qualidade do solo, com vistas ao aumento da produção,por exemplo. Por outro lado, com adições acima de 6 Mg ha-1 de palha houve um aumento no estoque de carbono no solo, chegando a um incremento de 11,9% quando adicionados 8 Mg ha-1 de palha.
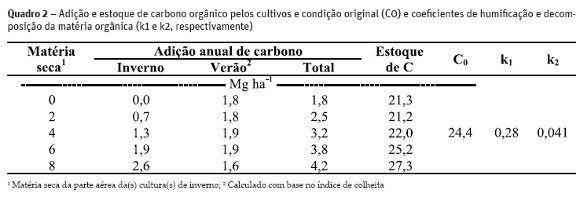
Esta dinâmica está relacionada com a velocidade dos processos de humificação (k1) e decomposição (k2) da matéria orgânica do solo, com valores de 0,28 e 0,041, respectivamente (Quadro 2). Assim, mesmo mantendo altas adições de resíduos, com alto k1, a sua incorporação no solo via transformações microbiológicas é acelerada, o que aumentaria os estoques de COS rapidamente. No entanto, com alto k2, mesmo mantendo altas taxas de incremento de palha, a adição líquida de carbono no solo acaba sendo baixa, em função da perda de COS pela degradação da matéria orgânica do solo.
O aporte de resíduos maior que 6 Mg ha-1 associado ao menor revolvimento do solo, resulta na retirada de CO2 da atmosfera e sua retenção no solo na forma de carbono orgânico. Utilizando as equações 1 e 2, e tomando por base uma área de um hectare, a combinação de manejos (6 e 8 Mg ha-1 de resíduos com as culturas de verão – Quadro 2) resultou em uma fixação de 3,9 e 4,3 Mg CO2 ano via fotossítes, respectivamente, enquanto o solo emitiu para a atmosfera, em 2000, 3,7 Mg de CO2, resultando em um influxo líquido de 0,2 e 0,6 Mg CO ha-1ano-1, respectivamente. Embora este influxo seja pequeno comparado a 1,79 Mg CO2 ha-1ano-1, como reportado por Bayer et al. (2000) que avaliou um sistema de plantio direto com cobertura de solo composta de aveia+vica/milho+caupi, os valores evidenciam a necessidade de adição e manutenção de altas quantidades de resíduos sobre o solo. Isso ocorre especialmente quando a cobertura de inverno não é cultivada em consórcio, para aumentar o teor de matéria orgânica.
O valor calculado de k1 está acima de 7,7 e 23% reportados na literatura (Bolinder et al., 1999; Gregorich et al., 1995). Isso pode estar associado a três fatores principais: i) a camada amostrada de solo (0,10 m), uma vez que a atividade microbiológica é mais intensa quanto mais próxima a superfície (maior temperatura, oxigenação e material orgânico); ii) a qualidade morfológica e física do resíduo adicionado, sendo que a decomposição varia entre gramíneas ou leguminosas, e se os resíduos são triturados pela colheita mecânica, pois aumenta a área superficial específica que facilita a decomposição; e iii) a quantidade de carbono alocado pelas raízes, uma vez que a quantidade de raízes foi estimada (30%) com base na produção de matéria seca da parte aérea. Diferente do carbono adicionado via resíduos da parte aérea, o carbono alocado pelas raízes é adicionado diretamente no perfil do solo, reduzindo perdas por oxidação quando da transformação do resíduo em matéria orgânica no solo, bem com o material está protegido fisicamente do ataque de microrganismos, demorando mais para ser decomposto, conforme apontam estudos de Rasse et al. (2005).
A taxa de perda da matéria orgânica do solo (k2) foi de 4,1% sendo considerada alta quando comparada a dados de Bayer et al., (2000) que encontraram valor de k2 para plantio direto, em mesmo solo, de 2,9%. Considerando que as perdas de matéria orgânica por erosão foram mínimas durante o período de condução do experimento, atribui-se isso principalmente a menor camada de solo avaliada (0,10 m comparada a 0,175 m de Bayer et al., 2000 e 2006b), a menor proteção física da matéria orgânica pelos agregados (Bayer, 1996), a menor razão C:N e lignina:N (baixa entrada de nitrogênio via leguminosas, principalmente), a menor associação organo-mineral, e a localização da MOS no perfil do solo (Jastrow e Miller, 1998).
Respiração microbiana basal
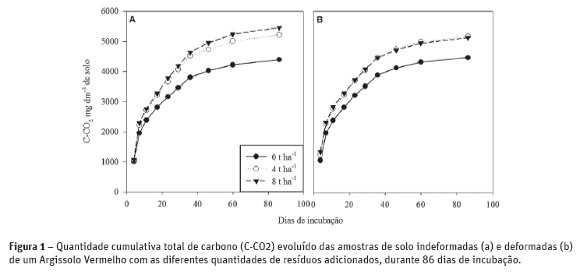
Após 86 dias de incubação verificou-se que não houve diferença significativa na respiração microbiana basal entre amostras deformadas e indeformadas (Quadro 3), dentro de cada tratamento independentemente da quantidade de resíduos adicionados. Porém, amostras de solo coletadas e mantidas indeformadas apresentaram diferenças significativas na evolução do C-CO2 emitido pela microbiota do solo para os diferentes aportes de resíduo, o que não foi verificado entre as amostras deformadas (Quadro 3). Tal comportamento pode ser creditado ao fato de que, as amostras deformadas foram desestruturadas e homogeneizadas enquanto que nas amostras indeformadas houve a preservação do ambiente em cada camada do perfil de solo dentro do tubo, o que pode ter permitido, pela conservação do microambiente, a maior taxa de emissão de CO2 nos tratamentos com maior aporte de resíduo.

Para as amostras indeformadas, o tratamento sem a adição de resíduo (0 Mg ha-1), apresentou menor respiração basal 4391,7 mg dm-3 comparado aos tratamentos com 4 e 8 Mg ha-1 que não distinguiram-se entre si, apresentando valores de 5318,7 e 5458,1 mg dm-3, respectivamente. Balota et al. (1998) cita que além de todas as variáveis de solo, manejo e clima, as condições metodológicas analíticas podem influenciar diretamente na taxa de C-CO2 emitido pela microbiota do solo.
Na Figura 1, é possível notar que as amostras indeformadas (Figura 1A) e deformadas (Figura 1B) demonstraram o comportamento de evolução de CO2 durante o período de incubação. Isto demonstra que o método de quantificação da respiração basal com amostras indeformadas pode ser uma estratégia importante em solos com sistema de plantio direto consolidado, onde não há a desestruturação do solo, impostas por preparos primários de solo como aração ou gradagens e aporte contínuo de resíduos. Segundo Follet e Schimel (1989) a maior liberação de CO2 geralmente é devida à maior atividade biológica que, por sua vez, está diretamente relacionada com a quantidade de carbono lábil existente no solo. Isso se refere diretamente a quantidade de material vegetal aportado, conforme evidenciado neste estudo para o tratamento com adição de 8 Mg ha-1 de resíduo. Entretanto, a interpretação dos resultados da atividade biológica deve ser feita com critério, uma vez que, em curto prazo, uma alta taxa de respiração pode significar liberação de nutrientes para as plantas e, em longo prazo, perda de carbono orgânico do solo para a atmosfera (Parkin et al., 1996). Estas informações reforçam a importância da adoção de sistemas de produção que priorizem rotação de culturas e manutenção contínua de altos aportes de resíduos vegetais na superfície do solo. Contudo, os valores de respiração basal descritos pela literatura são bastante variados sendo atribuídos a variabilidade de solo, clima e cobertura vegetal (Balota et al, 1998; Wardle, 1999; Hungria e Vargas, 2000; Hungria, 2009).
Carbono da biomassa microbiana (BMS-C) e quociente metabólico
No Quadro 4 são apresentados a média dos valores do BMS-C e do qCO2 para os tratamentos com adição de zero, 4 e 8 Mg ha-1 de resíduos. Foram observadas diferenças significativas para o BMS-C entre os tratamentos com adição de 4 e 8 Mg ha-1, com maior valor no último tratamento. Isso corrobora com o aumento na quantidade de matéria orgânica do solo (evidenciada pelo aumento nos teores de carbono – Quadro 2) que é fonte de nutrientes (Gnankambary et al., 2008) e energia para os microrganismos heterotróficos e plantas cultivadas (Bayer e Mielniczuk, 2008).
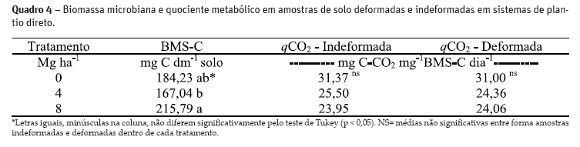
O tratamento com adição de 0 Mg ha-1 de resíduos apresentou valor absoluto de BMS-C intermediário, embora não tenha apresentado diferença significativa aos demais tratamentos. Isto pode ser um efeito momentâneo, causado por raízes e seus exsudatos, uma vez que a parcela estava sendo cultivada com aveia (a coleta das amostras de solo foi realizada em estágio reprodutivo desta cultura), porém não recebeu adição de resíduos da parte aérea, pois fora colhida e retirada da parcela. Assim, a menor quantidade de carbono no solo restringiria o crescimento da população mantendo um valor de respiração basal menor (Quadro 3). Com o aporte de carbono lábil via raízes e exsudatos, a população microbiana anteriormente sobre estresse, aumentaria momentaneamente, e após seria reduzida com o consumo desta fonte de carbono. Efeito semelhante foi encontrado e discutido por Balota et al, . (1998) e Franchini et al, . (2007).
A análise do quociente metabólico não identificou diferenças significativas entre tratamentos ou forma de coleta das amostras (Quadro 4). O quociente metabólico (qCO2) apresentou valores variando entre 23,95 e 31,37 mg de CO2 mg de BMS-C dia-1(Quadro 4). Apesar de não haver diferenças significativas entre os tratamentos, os quocientes metabólicos das duas formas de amostragem de solo mostram uma redução na quantidade de C-CO2 perdida à medida em que aumenta a dose de resíduo aplicado em superfície. O maior valor de quociente metabólico no tratamento zero é indicativo de que a condição de estresse alimentar faz com que os microrganismos gastem mais energia (consomem mais substrato) para manter a comunidade microbiana (Carneiro et al., 2008). Segundo Sparling (1997), quando a biomassa microbiana se torna mais eficiente, o que é indicado por um quociente metabólico menor, há maior proporção de C incorporado no tecido microbiano, explicando o maior valor de BMS-C encontrado no tratamento com a adição de 8 Mg ha-1 de resíduos corroborando com os resultados.
Conclusões
O estoque de carbono orgânico foi afetado pelo aporte de resíduos. A camada amostral de solo, características qualitativas dos resíduos e a proteção da matéria orgânica aparentam constituírem os fatores responsáveis pelos altos valores de k1 e k2 encontrados. O carbono da biomassa microbiana foi sensível ao incremento de resíduos, ao passo que o quociente metabólico não.
A utilização de amostras indeformadas para a análise da respiração basal, mostrou resultados semelhantes às amostras deformadas. Entretanto, amostras indeformadas apresentaram maior sensibilidade na identificação de variações entre tratamentos. Assim, outros estudos devem ser realizados buscando identificar e adequar esta metodologia de coleta.
Agradecimentos
Agradecemos ao CNPq e CAPES pelo apoio financeiro. À Estação Experimental Agronômica da Universidade Federal do Rio Grande do Sul pela concessão da área para coleta e aos professores e colaboradores.
Referências Bibliográficas
Anderson, T.H. e Domsch, K.H. (1993) The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil Biology and Biochemistry, vol.25, n.3, p. 393-395. [ Links ]
Balota, E.L.; Colozzi-Filho, A.; Andrade, D.S. e Hungria, M. (1998) Biomassa microbiana e sua atividade em solos sob diferentes sistemas de preparo e sucessão de culturas. Revista Brasileira de Ciência do Solo, vol.22, p. 641-649. [ Links ]
Bayer, C. (1996) Dinâmica da matéria orgânica em sistemas de manejo de solos. Tese de Doutoramento. Faculdade de Agronomia, Universidade Federal do Rio Grande do Sul, Porto Alegre. 240 p. [ Links ]
Bayer, C.; Mielniczuk, J. e Martin-Neto, L. (2000) Efeito de sistemas de preparo e de cultura na dinâmica da matéria orgânica e na mitigação das emissões de CO2. Revista Brasileira de Ciência do Solo, vol.24, p.599-607. [ Links ]
Bayer, C.; Martin-Neto, L.; Mielniczuk, J.; Pavinato, A. e Dieckow, J. (2006a ) Carbon sequestration in two Brazilian Cerrado soils under no-till. Soil and Tillage Research, vol.86, n.2, p. 237-245. [ Links ]
Bayer, C.; Lovato, T.; Dieckow, J.; Zanatta, J.A. e Mielniczuk, J. (2006b) A method for estimating coefficients of soil organic matter dynamics based on long-term experiments. Soil and Tillage Research, vol.91, n.1-2, p.217-226. [ Links ]
Bayer, C. e Mielniczuk, J. (2008) Dinâmica e função da matéria orgânica. In: Santos, G.A.; Silva, L.S.; Canellas, L.P. e Camargo, F.A.O. (Ed.) Fundamentos da matéria orgânica do solo: Ecossistemas tropicais e subtropicais, 2ª ed. Porto Alegre, Editora Metrópole, p. 7-18. [ Links ]
Bolinder, M.A.; Angers, D.A.; Gregorich, E.G. e Carter, M.R. (1999) The response of soil quality indicators to conservation management. Canadian Journal of Soil Science, vol.79, n.1, p. 37-45. [ Links ]
Carneiro, M.A.C.; Siqueira, J.O.; Moreira, F.M.S. e Soares, A.L.L. (2008) Carbono orgânico, nitrogênio total, biomassa e atividade microbiana do solo em duas cronossequências de reabilitação após a mineração de bauxita. Revista Brasileira de Ciência do Solo, vol.32, n.2, p. 621-632. [ Links ]
Chaer, G.M. e Tótola, M.R. (2007) Impacto do manejo de resíduos orgânicos durante a reforma de plantios de eucalipto sobre indicadores de qualidade de solo. Revista Brasileira de Ciência do Solo, vol.31, n.6, p. 1381-1396. [ Links ]
Dalal, R.C. e Mayer, R.J. (1986) Long-term trends in fertility of soils under continuous cultivation and cereal cropping in southern Queensland. II. Total organic carbon and its rate of loss from the soil profile. Australian Journal of Soil Research, vol.24, n.2, p. 281-292. [ Links ]
Follet, R.F.e Schimel, D.S. (1989) Effect of tillage practices on microbial biomass dynamics. Soil Science Society of America Journal, vol.53, n.4, p. 1091-1096. [ Links ]
Franchini, J.C.; Crispino, C.C.; Souza, R.A.; Torres, E. e Hungria, M. (2007) Microbiological parameters as indicators of soil quality under various soil management and crop rotation systems in Southern Brazil. Soil and Tillage Research, vol.92,n.1-2, p. 18-29. [ Links ]
Gnankambary, Z.; Ilstedt, U.; Nyberg, G.; Hien,V. e Malmer, A. (2008) Nitrogen and phosphorus limitation of soil microbial respiration in two tropical agroforestry parklands in the south Sudanese zone of Burkina Faso: The effects of tree canopy and fertilization. Soil Biology and Biochemistry, vol.40, n.2, p.350-359. [ Links ]
Gregorich, E.G.; Ellert, B.H. e Monreal, C.M. (1995)Turnover of soil organic matter and storage of corn residue carbon estimated from natural 13C abundance. Canadian Journal of Soil Science, vol.75, n.2, p. 161-167. [ Links ]
Hungria, M.; Franchini, J.C.; Brandão-Junior, O.; Kaschuk, G. e Souza, R.A. (2009) Soil microbial activity and crop sustainability in a long-term experiment with three soil-tillage and two crop-rotation systems. Applied Soil Ecology, vol.42, n.3, p. 288-296. [ Links ]
Hungria, M. e Vargas, M.A.T. (2000) Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Research, vol.65, n.2-3, p. 151-164. [ Links ]
Jastrow, J.D. e Miller, R.M. (1997) Soil aggregate stabilization and carbon sequestration: Feedbacks through organo-mineral associations. In: Lal, R.; Kimble, J.M., Follet, R.F. e Stewart, B.A. (Eds.)Soil processes and the carbon cycle. Boca Raton (etc.), USA, CRC Press, p.207-224. [ Links ]
Jenkinson, D.S. e Powlson, D.S. (1976) The effects of biocidal treatments on metabolism in soil. V: A method for measuring soil biomass. Soil Biology and Biochemistry, vol.8, n.3, p. 209-213. [ Links ]
Moreira, F.M.S. e Siqueira, J.O. (2006) – Microbiologia e bioquímica do solo. 2ª ed. Lavras, Brasil, Editora UFLA, 729p. [ Links ]
Nielsen, M.N. e Winding, A. (2002) – Microorganisms as indicators of soil health. Denmark, National Environmental Research Institute, 82p. (Technical Report n. 388) [ Links ]
Parkin, T.B.; Doran, J.W. e Francop-Vizcaíno, E. (1996) Field and laboratory tests of soil respiration. In: Doran, J.W. e Jones, A.J. (Ed.). Methods for assessing soil quality. Madison, USA, Soil Science Society of America, p.231-245. [ Links ]
Pereira, A.A.; Hungria, M.; Franchini, J.C.; Kaschuk, G.; Chueire, L.M.O.; Campo, R.J. e Torres, E. (2007) Variações qualitativas e quantitativas na microbiota do solo e na fixação biológica do nitrogênio sob diferentes manejos com soja. Revista Brasileira de Ciência do Solo, vol.31, n.6, p. 1397-1412. [ Links ]
Petry, H.B.; Oliveira, D.; Silveira, A.O.; Andreazza, R. e Camargo, F.A.O. (2012) Qualidade biológica do solo de sistemas de cultivo em pomares de Citrus deliciosa. Ciência e Natura, vol.34, n.1, p. 63-79. [ Links ]
Rasse, D.P.; Rumpel, C. e Dignac, M.-F. (2005) Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant and Soil, vol.269, n.12, p. 341-356. [ Links ]
Santos, H.P.; Tomm, G.O. e Kochhann, R.A. (2003) Rendimento de grãos de milho em função de diferentes sistemas de manejo de solo e de rotação de culturas. Revista Brasileira de Agrociência, vol.9, n.3, p. 251-256. [ Links ]
Segatto, M.P.; Andreazza, R.; Bortolon, L.; Santos, V.P.; Gianello, C. e Camargo, F.A.O. (2012) Decomposição de resíduos industriais no solo. Ciência e Natura, vol.34, n.1, p. 49-62. [ Links ]
Sequinatto, L. (2010) Qualidade física do solo e rendimento de grãos num Argissolo em processo de recuperação. Tese de Doutoramento. Universidade Federal do Rio Grande do Sul, Porto Alegre.141p. [ Links ]
Shukla, M. K.; Lal, R. e Ebinger, M. (2006) Determining soil quality indicators by factor analysis. Soil and Tillage Research, vol.87, n.2, p. 194-204. [ Links ]
Sparling, G. P. (1997) Soil microbial biomass, activity and nutrient cycling as indicators of soil health. In: Pankhurst, C.; Doube, B.M e Gupta, V.V.S.R. (Eds.) Biological indicators of soil health. Wallingford, CAB International, p.97-119. [ Links ]
Streck, E.V.; Kämpf, N.; Dalmolin, R.S.D.; Klamt, E.; Nascimento, P.C.; Schneider, P.; Giasson, E. e Pinto, L.F.S. (2008) Solos do Rio Grande do Sul. 2ª ed. Porto Alegre, EMATER/RS-ASCAR, 222p. [ Links ]
Trannin, I.C.B.; Siqueira, J.O. e Moreira, F.M.S.,(2007) Características biológicas do solo indicadoras de qualidade após dois anos de aplicação de biossólido industrial e cultivo de milho. Revista Brasileira de Ciência do Solo, vol.31, n.5, p.1173-1184. [ Links ]
Vieira, F.C.B. (2007) Estoques e labilidade da matéria orgânica e acidificação de um argissolo sob plantio direto afetados por sistemas de cultura e adubação nitrogenada. Tese de Doutoramento. Porto Alegre, Universidade Federal do Rio Grande do Sul, 123p. [ Links ]
Wardle, D.A. (1999) Is sampling effect a ´problem for experiments investigating biodiversity ecosystem function relationship? Oikos, vol.87, n.2, p. 403-407. [ Links ]
Zanatta, J.A.; Bayer, C.; Dieckow, J.; Vieira, F.C.B. e Mielniczuk, J. (2007) Soil organic carbon accumulation and carbon costs related to tillage, crop systems and nitrogen fertilization in a subtropical Acrisol. Soil and Tillage Research, vol.94, n.2, p. 510-519. [ Links ]
Recebido/Received: 2013.02.18
Aceitação/Accepted: 2013.03.03

