










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Ciências Agrárias
Print version ISSN 0871-018X
Rev. de Ciências Agrárias vol.36 no.2 Lisboa Apr. 2013
Lavandula stoechas subsp. luisieri and L. pedunculata - phytochemical study, micromorphology and histochemistry
Lavandula stoechas subsp. luisieri e L. pedunculata - estudo fitoquímico, micromorfologia e histoquímica
Generosa Teixeira1, Ana Isabel Correia2, Teresa Vasconcelos3, Daniela Feijão2 and Ana Margarida Madureira4
1 Faculdade de Farmácia, Centro de Biologia Ambiental, Universidade de Lisboa, Av. Prof. Gama Pinto, 1649-003, Lisboa, Portugal. E-mail: gteixeira@ff.ul.pt, author for correspondence
2 Faculdade de Ciências, Centro de Biologia Ambiental, Universidade de Lisboa, C2, Campo Grande, 1749-016 Lisboa, Portugal
3 Instituto Superior de Agronomia, Tapada da Ajuda, 1349-017 Lisboa, Portugal
4 Faculdade de Farmácia, iMed-UL, Universidade de Lisboa, Av. Prof. Gama Pinto, 1649-003, Lisboa, Portugal
ABSTRACT
Extracts of different polarities were obtained from plants of Lavandula stoechas subsp. luisieri and Lavandula pedunculata collected in the Center and Southwest Portugal. In order to have an idea on the possible classes of compounds present in each extract, they were subjected to a preliminary phytochemical screening through thin layer chromatography on silica gel plates, developed in appropriated mixtures of eluents and revealed with the specific reagents. Both taxa showed a hydrophilic and lipophilic chemical composition, being more diverse in L. stoechas subsp. luisieri. The micromorphology and histochemistry of secretory structures were also investigated by light and scanning electron microscopy. The common types of glandular trichomes, found in the Lamiaceae, peltate and capitate type I and II, were identified in each species as well as non-glandular stellate trichomes. In addition to these, L. pedunculata also presented capitate type III trichomes and still more two types of secretory trichomes: i) stellate type with both glandular and non glandular arms and ii) another type of capitate trichome, with a long and broad stalk formed by 2 cells, the basal is conical and much larger than the second, this similar to a ring, which supports a pluricelular head. These types of trichomes appear in the leaves but predominate in the flowers. The histochemical tests confirmed the existence of hydrophilic and lipophilic metabolites in glandular trichomes, except in the peltate hairs of the leaves of L. stoechas subsp. luisieri where only lipophilic secretions prevail.
Keywords: Lavandula, trichomes, phytochemical study, micromorphology, histochemistry
RESUMO
Extratos de diferentes polaridades foram obtidos de plantas de Lavandula stoechas subsp. luisieri e L. pedunculata, colhidas no centro e sudoeste de Portugal. Para ter uma ideia sobre as possíveis classes de compostos presentes em cada extracto, estes foram submetidos a um estudo fitoquímico preliminar através de cromatografia em camada fina, em placas de sílica gel, desenvolvidos em misturas apropriadas de eluentes e reveladas com reagentes específicos. Os dois taxa apresentaram uma composição química de natureza hidrofílica e lipofílica, sendo mais diversificada a encontrada em Lavandula stoechas subsp. luisieri.
A micromorfologia e histoquímica das estruturas secretoras foram observadas através de microscopia ótica e eletrónica de varrimento. Os tipos comuns de tricomas glandulares encontrados nas Lamiaceae, tricomas peltados e capitados tipo I e II, foram identificados nos 2 taxa, assim como tricomas não-glandulares de tipo estrelado. Além destes, L. pedunculata apresentou ainda tricomas capitados tipo III e mais dois tipos de tricomas secretores: i) estrelado com braços glandulares ou estrelado com braços não glandulares e glandulares e ii) um outro tipo de tricoma capitado, de pedúnculo longo e largo, formado por 2 células, a basal cónica, muito maior do que a 2ª célula, esta semelhante a um anel, que suporta uma cabeça pluricelular. Estes dois tipos de tricomas aparecem em folhas mas predominam nas flores. Testes histoquímicos confirmaram a existência de metabolitos secundários de natureza hidrofílica e lipofílica nos tricomas glandulares, salvo nos tricomas peltados das folhas de L. luisieri subsp. stoechas, onde predominaram apenas secreções lipofílicas.
Palavras chave: Lavandula, tricomas, estudo fitoquímico, micromorfologia, histoquímica
Introduction
The Lamiaceae is one of the most important botanical families found in the area of Mediterranean influence. Due to medicinal properties and economic potential in the food and fragrance industries, there is a great interest in their plant species biological and chemical characterization. Nevertheless, only Lavandula species of considerable economic significance are usually well documented (Cavanagh, 2002). In contrast, the native taxa have received much less attention and are only reported as aromatic species and little information is available on their glandular structures, their chemicals and their activities (Chizzola, 2006; Combrinck et al., 2007). This is the case of some Lavandula species and subspecies from Portugal, among them Lavandula stoechas subsp. luisieri (Rozeira) Rozeira (Syn: L. luisieri; L. stoechas var. luisieri) and Lavandula pedunculata (Mill.) Cav. (Syn: L. pedunculata subsp. sampaiana; L. stoechas subsp. pedunculata) (Morales, 2010).
Some morphological characters and the chemical composition are known to be affected by environmental conditions and by the phenological state of the plants. These are two approaches for plant description and classification also applied to the Lamiaceae where polymorphisms and chemiotypes are known (Husain et al., 1990; Harley et al. 2004; Salmaki et al., 2009). The great majority of the members of this family show non glandular and glandular trichomes, located in the epidermis of aerial parts which may vary in their density and morphology. According to the structure and mode of secretion, glandular trichomes can be classified into two general types, peltate and capitate (Werker et al., 1985). The last are morphologically more diverse and can be divided into other types, also according to its structure and also to its method of secretion (Werker et al., 1985).
In order to achieve a better knowledge about the chemical composition and micromorphology of the Portuguese taxa previously mentioned, we performed a series of increasing polarity extracts and carried out their preliminary phytochemical screening by chromatographic methods. Light and scanning electron microscopy were used to elucidate about the secretory structures and histochemical tests were done to achieve cell and glandular localization of the main chemical groups.
Material and Methods
Plant material
Small branches of both species at flowering stage were collected between 2008 and 2011, from plants growing in open field, in the Center and Southwestern regions of Portugal. Vouchers were deposited at the Lisbon Botanical Garden Herbarium (LISU) and the Lisbon Agronomic Institute Herbarium (LISI), (Table 1). For obtaining plant extracts we used leaves and flowers of only 2 taxa (Table 1). For micromorphological and histochemical studies full-expanded leaves and flowers were considered.
Preparation of crude plants extracts
Crude plants extracts were prepared by extracting sequentially 100 g of dried powdered plant material with 300 ml of n-hexane, dichloromethane, ethyl acetate, methanol and water, during 24-h, at room temperature with occasional shaking. After filtration, the extracts were concentrated under reduced pressure at 40-45 ºC and stored at 4 ºC until use.
Phytochemical screening
Phytochemical analysis was carried out through thin layer chromatography on silica gel plates developed with appropriated mixtures of eluents. Spots were revealed with the following spray-reagents: Dragendorff reagent for alkaloids, anisaldehyde-sulfuric acid reagent for terpenes, Natural Products-Polyethylene Glycol (NEU) reagent for flavonoids and Fast Blue salt reagent for phenolic compounds, prepared according to Wagner and Blader (1996). Results were displayed semi-quantitatively in a range between absence (-) and strongly present (+++).
Light microscopy (LM)
Samples of fresh, fixed and paraffin embedded material were used. The fresh material was hand-sectioned transversely and paradermally clarified with sodium hypochlorite and washed in distilled water. Fixation was achieved in a 2.5% glutaraldehyde solution in a 0.1M sodium phosphate buffer, pH 7.2, for 5 h at 4°C and dehydrated in a graded ethanol series (Hayat, 1981). Some fixed material was embedded in paraffin (Ruzin, 1999) and tissue blocks were sectioned at 10-12 µm using a Leitz 1512 Minot microtome.
A Nikon Labophot 2 microscope was used and images were obtained with a Nikon FX-35W camera. Characters were observed on fifteen different observations per specimen.
Scanning electron microscopy (SEM)
Plant material was fixed as above, dried with liquid CO2 at a critical point, on a Critical Point Polaron BioRad E3500, and coated with gold on a Jeol JFC-1200. Samples were examined at 15 kV with a Jeol JSM-5220 LV scanning electron microscope. Measurements and counts were achieved on SEM micrographs obtained by computer assisted image analysis.
Histochemistry
The following histochemical tests were carried out: (a) Sudan Red for total lipids (Johansen, 1940); (b) Nile Blue procedure for neutral and acidic lipids (Jensen, 1962); (c) Copper acetate / Rubeanic acid for fatty acids (Ganter and G. Jolés, 1969); (d) Nadi reagent for essential oils and oleoresins detection (David and Carde, 1964); (e) 2,4-dinitrophenilhidrazine for terpenes with a carbonyl group (Ganter and G. Jolés, 1969); (f) Antimony trichloride for steroids (Hardman and Sofora, 1972) (g) Ferric trichloride for phenols (Johansen, 1940); (h) Vanillin hydrochloric for tannins (Johansen, 1940); (i) PAS for total polysaccharides (Feder and O`Brien (1968); (j) Ruthenium Red for pectins (Jensen, 1962) ; (k) Tannic acid for mucilages (Pizzolato and Lillie, 1973) and (l) Dittmar reagent for alkaloids (Johansen, 1940). Standard control procedures were carried out at the same time. Observations were done in the mentioned LM and results were semi quantitatively analised, between negative (-) and strongly positive (+++).
Results
Phytochemical screening
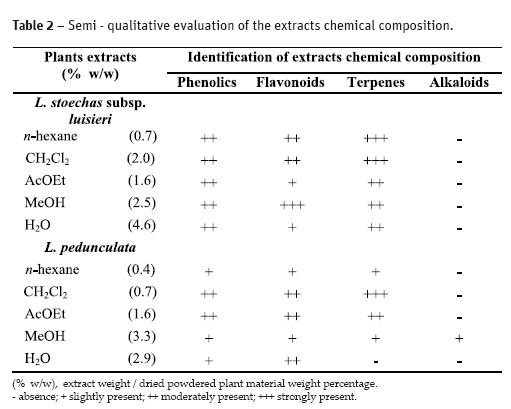
The results of the phytochemical screening and the yield of the dried extracts (as w/w percentage of the starting dried material) are presented on Table 2. It is clear that all plant extracts were negative to alkaloids, all except the methanol extract of L. pedunculata. The extracts obtained seemed to be rich in phenolics, flavonoids and terpenes. L. stoechas subsp. luisieri extracts seemed to have a higher content and a higher variety of terpenic compounds.
Plant trichomes
L. stoechas subsp. luisieri and L. pedunculata bear an indumentum in vegetative and flowering structures that includes non-glandular and glandular trichomes random distributed on both surfaces. The different populations maintain the characters of each species, only presenting some changes in the indumentum density. Both species showed short uni- and bicellular unbranched trichomes and mainly pluricellular branched trichomes of the stellate type. In the leaf upper epidermis the density of the last type of trichomes was smaller and they were also less long. In L. stoechas subsp. luisieri their stalk was up to 52-85 mm long and in L. pedunculata 60-128 mm and in both taxa they support 3-7 arms (Fig. 1A). In the lower epidermis this type of trichomes was denser and shorter, up to 26-39 mm in L. stoechas subsp. luisieri and 27-60 mm in L. pedunculata. In the flowers, only the fertile bracts and the sepals showed stellate trichomes and their stalk could reach up to 300 mm long. The petals did not have this kind of hairs, only uni- and pluricellular unbranched trichomes.
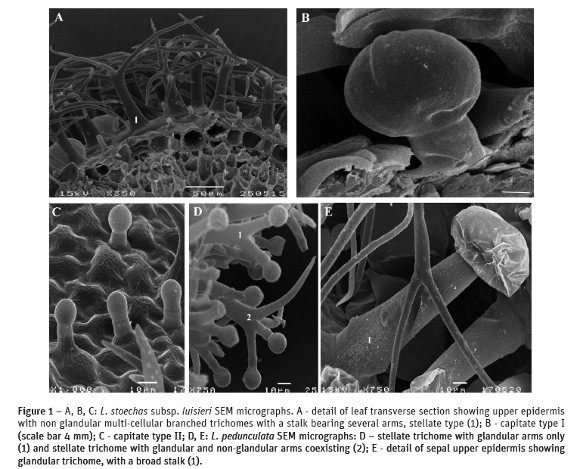
Concerning the glandular trichomes found, they belong to the peltate and capitate types. The peltate trichomes had 1 basal cell supporting 1 short stalk cell which bears an head composed of 8-12 glandular cells topped by a subcuticular space where the secretion products were stored. The capitate trichomes were much more variable in their morphology and different types could be distinguished: in L. stoechas subsp. luisieri, capitate type I with 1 basal cell, 1stalk cell and a head with 1-2 broad cells (Fig. 1B); capitate type II with 1 conical basal cell, 1-2 stalk cells and a round apical head (Fig. 1C). In the fertile bracts and sepals of L. pedunculata and in much smaller number in the leaves, the variety of secretor hairs was bigger: besides the peltate and capitate type I and II just described, there were also capitate type III trichomes. These showed 1 conical basal cell, 2-5 elongated stalk cells and a round head cell that might become cup-shaped at maturity (Fig. 3B; 3D). In this species besides the previous types of non-glandular and glandular trichomes, there were found other kinds of glandular hairs: stellate trichomes with 2-4 arms, similar in their measures and general structure to the non-glandular previous described, but their arms support glandular cells at the end, or these coexist with non-glandular end cells (Fig. 1D). In L. pedunculata we have found yet another type of secretor trichome, also distributed mainly in the fertile bracts and in the sepals, even if outnumbered. They showed a long and broad stalk, consisting of 2 cells, one conical elongated and another much smaller, similar to a ring, which supports a pluricellular head, and whose cells were covered by a cuticle (Fig. 1E). The petals of L. pedunculata did not show any of these kinds of trichomes.
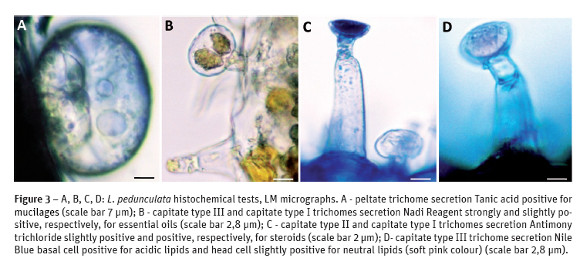
Histochemistry
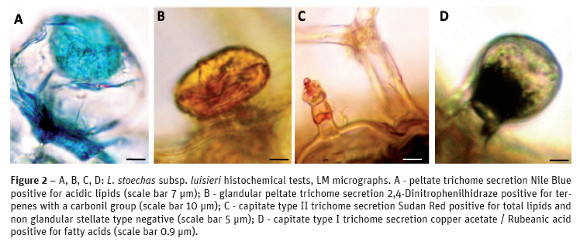
The results of the histochemical tests for the various types of glandular trichomes found in leaves and flowers of both taxa are shown in Table 3. The secreted compounds in the peltate trichomes were accumulated beneath the cuticular sheet which at maturity lifts to form a large sub-cuticular chamber for the secretory materials storage. Our observations did not reveal any pores or cracks through which the secretory material could exude. In L. stoechas subsp. luisieri specimens peltate trichomes stained slightly positive with Sudan Red and Rubeanic acid, stained positive with Nile Blue (Fig. 2A) and 2,4 - Dinitrofenilhidrazine (Fig. 2B), stained strongly positive with Nadi reagent and Antimony trichloride and stained negative with Vanillin hydrochloric, Ferric trichloride, PAS, Ruthenium Red and Dittmar reagent and the result was inconclusive with Tannic acid. In this subspecies the secretions of capitate trichomes, type I and II, found in the leaves are similar, the only difference is the staining intensity: they stained positive with Sudan Red (Fig. 2C), Nile Blue, Rubeanic acid (Fig. 2D), Antimony trichloride and Ferric trichloride, and stained negative with the remaining tests. In the flowers of L. stoechas subsp. luisieri specimens peltate and capitate trichomes stained slightly positive with PAS, Ruthenium Red and Tannic acid.
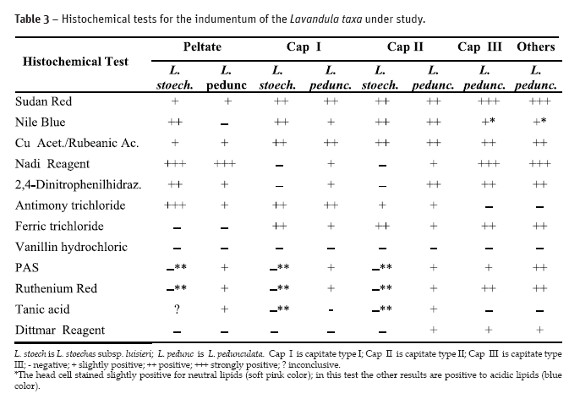
In L. pedunculata the histochemical tests concerning the peltate trichomes, from the leaves and the flowers, are negative with Nile Blue, Ferric trichloride, Vanillin, Dittmar Reagent and they are positive, although some intensity differences, with the remaining tests, including with Tannic acid (Fig. 3A). The tests of all types of capitate trichomes are also similar on their positive results, although some intensity differences (Fig. 3B and Fig. 3C). With the Nile Blue test the capitate type III trichome basal cell stained positive for acidic lipids and the head cell stained slightly positive, a soft pink color, for neutral lipids (Fig. 3D).
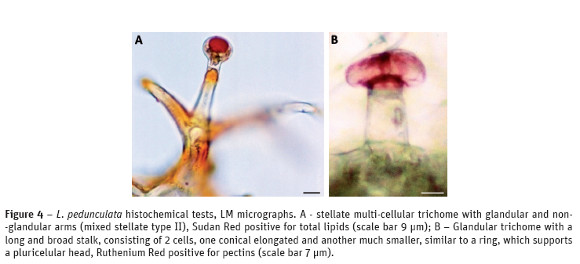
The results of the histochemical tests of the other kinds of trichomes found in L. pedunculata, stellate trichomes with glandular and non-glandular arms or only with glandular arms and in the other capitate type with a long and broad stalk, are similar to those found in capitate type III trichomes. The mode of secretion and accumulation of the secreted compounds in these hairs, including in the capitate type III, was beneath the cuticular sheet in a subcuticular chamber (Fig. 4A and Fig. 4B).
Discussion
The phytochemical analysis in the several extracts of the studied plants, points to a similar content of phenolics and flavonoids but the terpenic content is different. In L. stoechas subsp. luisieri extracts the number of terpenic compounds was higher and richer. Several authors mentioned that some necrodane derivatives characterize its essential oils (Feijão, 2011; González-Coloma et al., 2006; Matos et al., 2009; Baldovini et al., 2005; Sanz et al., 2004; Garcia-Vallejo et al., 1994), which seem to be absent in the remaining Lavandula spp. essential oils. Several authors working on Spanish material of L. stoechas mention the difficult differentiation of subspecies and varieties within this species due to high polymorphism, but they consider some chemiotypes based only on chemical data (Garcia-Vallejo et al., 1994). The essential oils of L. pedunculata are rich in fenchone (Zuzarte et al., 2009). In a previous work with specimens harvested in our locations, Feijão (2011) also confirmed the dominance of fenchone in the chemical composition of the essential oils of the same species.
L. stoechas subsp. luisieri and L. pedunculata showed non-glandular trichomes, small uni- and bicellular trichomes and abundant stellate trichomes. These hairs have a similar distribution in the leaves fertile bracts and sepals surfaces but stellate trichomes are not found in the petals. Based on their structure, glandular trichomes are generally classified as either peltate or capitate (Werker et al., 1985). Peltate trichomes do not present any diversity in their morphology; they are similar to those found in other Lamiaceae. The capitate trichomes vary greatly in structure and size. According to our observations L. pedunculata showed a wider diversity of secretory trichomes, such as the multicellular stellate trichomes which combine glandular and non-glandular arms. Zuzarte et al. (2010) also mentioned this kind of trichomes in samples of L. pedunculata from central north Portugal as well as in in vitro leaf cultures but no reference is done to the capitate type III and to the other type of capitate trichomes, those with the long and broad stalk with the pluricellular head that we found distributed either by the leaves or by the different floral parts but once again not in the petals. This kind of glandular trichomes is mentioned by Doaogey (1992) in leaves and flowers of L. stricta and L. coronopifolia, exemplars from the south of Saudi Arabia.
It should be noted that in the flowers of both taxa the petals showed little diversity of secretory trichomes. Only peltate hairs were seen and the greatest diversity and amount of trichomes arises in the fertile bracts and the sepals.
Data from histochemical tests showed that the metabolites accumulated in the glandular trichomes of the studied taxa have a distinct composition and that the secreted materials are complex in their nature. These tests contribute to a better understanding of the location of metabolites in cells and glandular structures. In the peltate hairs of L. stoechas subsp. luisieri leaves, the secreted compounds accumulated beneath the cuticule were most lipophilic, while in L. pedunculata leaves and flowers structuresthe presence of polysaccharides, mucilages and pectines were also revealed with PAS, Tannic acid and ruthenium red histochemical tests, respectively. In L. stoechas subsp. luisieri a slightly presence of these hydrophilic compounds was noticed in the fertile bracts and the sepals. The presence of these compounds in the glandular trichomes of other Lamiaceae has been reported by several authors (Werker, 1993; Corsi and Bottega, 1999; Ascensão et al., 1999; Marin et al., 2010; Rodrigues et al., 2013) but their function is not completely explained (Werker, 1993; Corsi and Bottega, 1999).
The histochemical results indicate that capitate trichomes of L. pedunculata produced a much more heterogeneous secretion than those of L. stoechas subsp. luisieri. Indeed capitate trichomes seem to contain a much more complex mixture of secretory products, containing several lipophilic compounds in addition to phenols, polysaccharides and traces of alkaloids. Similar contents have also been reported in other related genus of the Lamiaceae (Serrato-Valenti et al., 1997; Ascenção et al., 1999; Rodrigues et al., 2008; Rodrigues et al., 2013). It is accepted that some of these secondary metabolites have important ecological effects and provide plants with physical and chemical protection against herbivores and pathogens (Werker et al., 1985; Fahn, 1988; Harborne, 1997).
In L. pedunculata the other types of trichomes also have a mixture of lipophilic and hydrophilic compounds, very similar to those found in capitate type III trichomes. It is important to note that in the trichomes with glandular and non-glandular arms, both of them seem to accumulate secretions (Fig. 4A). Although their structure is quite different these trichomes seem to be more close to glandular trichomes in their secretory material and in their accumulation processes: beneath the cuticular sheet in a sub-cuticular chamber. The non-glandular hairs particularly the multi-cellular branched trichomes presumably assist glandular hairs in the mechanical defense and they are certainly also involved in preventing an excessive leaf transpiration.
Although the low specificity of some histochemical tests, they are widely used to locate metabolites in all kinds of glandular structures (Ascenção et al., 1999; Bisio et al., 1999; Combrick et al., 2007). These tests do not allow us to identify compounds just find the main chemical groups at the cellular level. In this sense the results from Table 2 and Table 3 show some similarity between species, concerning the presence of the main chemical groups, such as the alkaloids but also the phenolics and terpenes.
In conclusion, L. stoechas subsp. luisieri and L. pedunculata have different types of glandular trichomes and despite the need to continue the chemical characterization of each taxon, our phytochemical screening and the results of the histochemical tests point to a distinct chemical composition between them.
References
Ascensão, L.; Mota, L. and Castro, M. (1999) - Glandular trichomes on the leaves and flowers of Plectranthus ornatus: morphology, distribution and histochemistry. Annals of Botany,84: 437-447. [ Links ]
Baldovini, N.; Lavoine-Hanneguelle, S.; Ferrando, G.; Dusart, G. and Lizzani-Cuvelier, L. (2005) - Necrodane monoterpenoids from Lavandula luisieri. Phytochemistry, 66: 1651-1655. [ Links ]
Bisio, A.; Corallo, A.; Gastaldo, P.; Romussi, G.; Ciarallo, G. and Fontana, N. (1999) - Glandular hairs and secreted material in Salvia blepharophylla Brandegee ex Epling grown in Italy. Annals of Botany, 83: 441-452. [ Links ]
Cavanagh, H.M. and Wilkinson, J.M. (2002) - Review article: Biological activities of lavender essential oil. Phytotherapy Research, 16: 301-308. [ Links ]
Chizzola, R. (2006) - Volatile compounds from some wild growing aromatic herbs of the Lamiaceae from southern France. Plant Biosystems, 140: 206 -210. [ Links ]
Combrinck, S.; Plooy, G.; McCrindle, R. and Botha, B. (2007) - Morphology and Histochemistry of the Glandular Trichomes of Lippia scaberrima (Verbenaceae). Annals of Botany, 99: 1111-1119. [ Links ]
Corsi, G. and Bottega, S. (1999) - Glandular hairs of Salvia officinalis: new data on morphology, localization and histochemistry in relation to function. Annals of Botany, 84: 657-664. [ Links ]
David, J. and Carde, P. (1964) - Coloration différentielle des pseudophylles de Pin maritime au moyen réactif de Nadi. Comptes Rendus de l' Academie des Sciences Paris, Serie D, 258: 1338-1340. [ Links ]
Doaogey. A.R. (1992) - Types and occurrence of trichomes in two species of the genus Lavandula: L. stricta and L. coronopifolia. Pakistan Journal of Botany, 24: 131-141. [ Links ]
Fahn A. (1988) - Secretory tissues in vascular plants. New Phytologist, 108: 229-257. [ Links ]
Feder, N. and OBrien, T.P. (1968) - Plant microtechnique: some principles and new methods. American Journal of Botany, 55:123-42. [ Links ]
Feijão, D.M. (2011) - A Flora Medicinal e Aromática da Herdade da Ribeira Abaixo, Grândola (Estação de Campo, CBA): caracterização micromorfológica e dos óleos essenciais de Lavandula luisieri. Dissertação de Mestrado, FCUL. 58 p. [ Links ]
Ganter P. and Jolés G. (1969) - Histochimie normale et pathologique. Paris: Gauthier-Villars, 904 p. [ Links ]
Garcia-Vallejo, M.I.; Garcia-Vallejo, M.C.; Sanz, J.; Bernabé, M. and Velasco-Negueruela. (1994) - Necrodanes derivatives in Lavandula luisieri, new compounds to the plant kingdom. Phytochemistry, 36: 43-45. [ Links ]
González-Coloma, A.; Martín-Benito, D.; Mohamed, N.; García-Vallejo, M.C. and Soria, A.C. (2006) - Antifeedant effects and chemical composition of essential oils from different populations of Lavandula luisieri L. Biochemical Systematics and Ecology, 34: 609-616. [ Links ]
Harborne, J.B. (1997) - Plant secondary metabolism. In: Crawley MJ, eds. Plant ecology. 2nd ed. Berlin: Blackwell Publisher, 355 p. [ Links ]
Hardman, R. and Sofowora, E.A. (1972) - Antimony trichloride as a test for steroids in plant tissues. Stain Technology, 47: 205-208. [ Links ]
Harley, R.M.; Atkins, S.; Budantsev, A.L.; Cantino, P.D.; Conn, B.J.; Grayer, R.; Harley, M.M.; de Kok, R.; Krestovskaja, T.; Morales, R.; Pato, A.J.;Ryding, O. and Upson, T. (2004) –Labiatae, 167-282. In Kadereit, J.W. (ed.) The families and genera of vascular plants. Vol. VII, Lamiales. Berlin: Springer, 478 p. [ Links ]
Hayat, M. (1981) - Principles and techniques of electron microscopy: biological applications. 2nd ed London: Edward Arnold Publication, Ltd., 234 p. [ Links ]
Husain, S.Z.; Marin, D.P.; Silic, C.; Qaiser, M. and Petkovic, B. (1990) - A micromorphology study of some representative genera in the tribe Saturejeae (Lamiaceae). Botanical Journal Linnaean Society, 103: 59–80. [ Links ]
Jensen, W.A. (1962) - Botanical Histochemistry: principles and practice. San Francisco, Freeman, 408 p. [ Links ]
Johansen, D.A. (1940) - Plant microtechnique. New York, McGraw-Hill, 523 p. [ Links ]
Marin, M.; Jasnic, N.; Lakusic, D.; Duletic-Lausevic, S. and Ascensão, L. (2010) - The micromorphological Histochemical and Confocal Analysis of Satureja subspicata Bartl. Ex Vis. Glandular Trichomes. Archives of Biological Sciences, 62: 1143-1149. [ Links ]
Matos, F.; Miguel, M.G.; Duarte, J.; Venâncio, F.; Monteiro, C.; Correia, A.I.; Figueiredo, A.C.; Barroso, J.G. and Pedro, L.G. (2009) - Antioxidant capacity of the essential oils from Lavandula luisieri, L. stoechas ssp. lusitanica, L. stoechas ssp. lusitanica x L. luisieri and L. viridis grown in Algarve (Portugal). Journal of Essential Oil Research, 21: 327-336. [ Links ]
Morales, R . (2010) - Lavandula. Flora Iberica, vol XII. Real Jardín Botánico CSIC, Madrid. [ Links ]
Pizzolato, P. and Lillie, R.D. (1973) - Mayer´s tannic acid –ferric chloride stain for mucins. Journal Histochemistry and Cytochemistry, 21: 56-64. [ Links ]
Rodrigues, L.; Monteiro, P.; Póvoa, O.; Teixeira, G.; Moldão, M.; Figueiredo, A. and Monteiro, A. (2008) - Morphology of secretory structures and essential oil composition in Mentha cervina L. from Portugal. Flavour Fragrance Journal, 23: 340-347. [ Links ]
Rodrigues, L.; Póvoa, O.; Teixeira, G.; Figueiredo, A.C.; Moldão, M. and Monteiro, A. (2013) - Trichomes micromorphology and essential oil variation at different developmental stages of cultivated and wild growing Mentha pulegium L. populations from Portugal. Industrial Crops and Products, 43: 692–700. [ Links ]
Ruzin, S. (1999) - Plant microtechnique and microscopy. London: Oxford University Press, 336 p. [ Links ]
Salmaki, Y.; Zarre, S.; Jamzad, Z. and Brauchler, C. (2009) - Trichome micromorphology of Iranian Stachys (Lamiaceae) with emphasis on its systematic implication. Flora, 204: 371-381. [ Links ]
Sanz, J.; Soria, A.C. and García-Vallejo M.C. (2004) - Analysis of volatile components of Lavandula luisieri L. by direct thermal desorption-gas chromatography-mass spectrometry. Journal of Chromatography, 1024: 139-146. [ Links ]
Serrato-Valenti, G.; Bisio, A.; Cornara, L. and Ciarallo, G. (1997) - Structural and histochemical investigation of the glandular trichomes of Salvia aurea L. leaves and chemical analysis of the essential oil. Annals of Botany, 79: 329-336. [ Links ]
Wagner, H. and Blader, S. (1996) - Plant drug analysis: A Thin Layer Chromatography Atlas. 2nd ed. Berlin: Springer-Verlag, 384 p. [ Links ]
Werker, E.; Ravid, U. and Putievsky, E. (1985) -Structure of glandular hairs and identification of the main components of their secreted material in some species of the Labiatae. Israel Journal of Botany, 34: 31-45. [ Links ]
Werker, E. (1993) - Function of essential oil secreting glandular hairs in aromatic plants of the Lamiaceae: a review. Flavour and Flagrance Journal, 8: 249-255. [ Links ]
Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Dinis, A.M.; Canhoto, A.M. and Salgueiro, L. (2009) - Chemical Composition and Antifungal Activity of the Essential Oils of Lavandula pedunculata (Miller) Cav. Chemical Biodiversity, 6: 1283-1292. [ Links ]
Zuzarte, M.; Dinis A.M.; Cavaleiro, C.; Salgueiro, L. and Canhoto, J.M. (2010) - Trichomes, essential oils and in vitro propagation of Lavandula pedunculata (Lamiaceae). Industrial Crops Products, 32: 580-587. [ Links ]
Recebido/Received: 2012.02.28
Aceitação/Accepted: 2013.04.10

