










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.34 no.2 Lisboa jul. 2011
Potencialidade das leguminosas forrageiras anuais como fonte de azoto em agricultura biológica
Adelaide Perdigão1, João Coutinho2 e Nuno Moreira3
1ESAV – ISPV – Quinta da Alagoa Estrada de Nelas 3500 Viseu aperdigao25@gmail.com;
2ECVA – UTAD – Vila Real;
3ECAV – UTAD - Vila Real
RESUMO
A reduzida investigação, à escala mundial, sobre a utilização de novas espécies de leguminosas forrageiras em condições mediterrânicas como fonte de N, conduziu-nos ao presente estudo, que teve como objectivos avaliar a produção de matéria seca (MS), quantificar o N acumulado e determinar o potencial de fornecimento de N por diferentes plantas como culturas intercalares em duas datas de sementeira diferentes.
Foram utilizadas 6 leguminosas forrageiras: trevo balansa, trevo glandulífero, trevo vesiculoso, trevo encarnado, serradela vulgar e tremocilha; uma gramínea: azevém; uma consociação de azevém com trevo balansa e a vegetação espontânea. As sementeiras realizaram-se em Setembro e Outubro de 2007.
Os melhores resultados foram obtidos para a 2ª data de sementeira, para todas as variáveis. A serradela foi responsável pela maior produção de MS (7179 kg ha-1). O trevo balansa obteve o maior teor de N (26,85 g kg-1) e o maior potencial de fornecimento de N através da biomassa aérea (124 kg N ha-1).
Palavras – chave: adubação verde; agricultura biológica; azoto; leguminosas forrageiras anuais; sideração.
Potential nitrogen supply of annual forage legumes for organic farming in a mediterranean environment
ABSTRACT
The need of information about new species of annual forage legumes as green manure crops for Mediterranean environments, led us to this study with the objective of evaluating the nitrogen replacement value of these crops.
We evaluated the effects of six forage legumes: balansa clover, crimson clover, gland clover, arrowleaf clover, french serradella and yellow lupine; one grass (ryegrass); one mixture (ryegrass with balansa clover) e one control treatment (semi-natural vegetation) in two different sowing dates: September e October 2007.
Results on dry matter yield, nitrogen concentration e nitrogen uptake were obtained for these treatments in 2007/08. In most treatments the better results of dry matter yield, nitrogen concentration e nitrogen uptake/supply were found on the second sowing date. The french serradella had the highest yield of dry matter (7179 kg ha-1). Balansa clover had the highest value of nitrogen concentration (26.85 g kg-1) e nitrogen uptake (124 kg N ha-1).
Keywords: catch crops; annual forage legumes; green manure; nitrogen organic farming;
INTRODUÇÃO
As questões ambientais têm cada vez maior relevância e surge a necessidade de, em todas as áreas, se evitar ao máximo as agressões ao ambiente.
A agricultura não é excepção, e nas últimas décadas, o impacto das actividades agrícolas no ambiente tem tido especial destaque (Korsaeth, 2008), pelo que se acentua a necessidade de desenvolver práticas e formas de agricultura mais "amigas do ambiente", nomeadamente com menor aplicação de produtos de síntese e em que se favoreça a utilização dos recursos naturais, sem nunca esquecer a manutenção da rentabilidade das explorações. Estas preocupações ambientais têm estimulado a procura de fontes alternativas de N, que possibilitem a substituição total ou parcial dos fertilizantes minerais. A agricultura biológica é sugerida como uma alternativa para manter a fertilidade dos solos e reduzir os impactos ambientais negativos (Bergström e Kirchmann, 2004): utiliza os recursos naturais disponíveis, não utiliza produtos de síntese e mantém a rentabilidade da exploração.
Um dos principais obstáculos em agricultura biológica é a limitação do uso de fertilizantes azotados inorgânicos (Fowler et al., 2004), particularmente nas situações em que os solos têm pouco N disponível, como frequentemente sucede nas condições da agricultura mediterrânica.
O N é o nutriente que mais limita a produção de culturas, nomeadamente em agricultura biológica (Askegaard e Eriksen, 2008; Kirchmann et al., 2007). Este só pode ser fornecido através da aplicação de estrumes ou resíduos/subprodutos de culturas compostados, de adubos orgânicos e da utilização de culturas intercalares que disponibilizam N ao solo (Cavigelli et al., 2008; Olesen et al., 2007), sendo a fixação biológica do N (utilização de leguminosas) a principal fonte de N em agricultura biológica (Pietsch et al., 2007; Möller et al., 2008), pois a utilização de estrumes é restrita (Askegaard e Eriksen, 2007) e a utilização de adubos orgânicos têm um custo elevado e uma eficiência questionável na nutrição das culturas (Rodrigues et al., 2004)
A utilização de culturas intercalares, em agricultura biológica, é uma ferramenta importante na gestão do N (Möller et al., 2008), pois, através das suas capacidades de fixação biológica ou retenção de N, é considerada a principal fonte de N (Fowler et al., 2004; Goh et al., 2001). As culturas intercalares são culturas que entram em rotação com a cultura principal. Dentro destas podemos ter culturas com duas finalidades diferentes: apenas revestir o solo para diminuição da erosão, e essas serão designadas culturas de revestimento ou de cobertura, e as culturas para melhorar os níveis de fertilidade do solo (através da sua incorporação no solo), as designadas culturas para sideração. Dentro das culturas para sideração ainda podemos diferenciar a adubação verde, constituída por culturas que fixam o N atmosférico e o disponibilizam ao solo, aumentado desta forma os níveis de N no solo, e as culturas retentoras de N ("catch crops"), culturas que retêm o N do solo e o transferem à cultura seguinte, minimizando desta forma as perdas de N por lixiviação (Askegaard e Eriksen, 2007; Sainju et al., 2007, Sullivan, 2003; Thorup-Kristensen et al., 2003). Estas culturas são reconhecidas como fonte de N, sempre que fornecem N suficiente à cultura principal de forma sincronizada, ou seja, a libertação de N é gradual, de forma a compensar as necessidades da cultura principal (Bergström e Kirchmann, 2004).
Estudos efectuados por Sainju e Singh (2008) demonstraram que a introdução de culturas para sideração, quando comparada com a vegetação espontânea, aumenta o teor do N total do solo. Este aumento é maior com as leguminosas estremes e com a consociação entre leguminosas e gramíneas do que com as gramíneas estremes ou com a vegetação espontânea.
Nem todas as espécies de leguminosas forrageiras têm o mesmo potencial de produção de biomassa e fornecimento de N, sendo necessário identificar as espécies com maior potencial (Snapp et al., 2005) e determinar se obtêm melhores resultados isoladas ou em consociação com gramíneas.
A quantidade de N acumulado pelas culturas é maior nas leguminosas quando comparado com as não leguminosas e com a consociação entre leguminosas e não leguminosas (Fowler et al., 2004). Em estudos desenvolvidos por Fowler et al., (2004) na Nova Zelândia (Lincoln University) obtiveram-se valores de 100, 162 e 126 kg N ha-1 acumulado em gramíneas, leguminosas e consociação, respectivamente.
A contribuição em N pelas leguminosas pode ser muito variável. O N disponibilizado pelas adubações verdes com leguminosas pode variar entre os 0 e os 159 kg N ha-1 (Oyer e Touchton, 1990; Reinbott et al., 2004).
A adaptação das leguminosas forrageiras a determinado local depende muito das condições climáticas e tipo de solo (Nichols et al., 2007; Thiessen Martens et al., 2001). As condições climáticas são o factor que mais limita a escolha da espécie (Cherr et al., 2006). Até há pouco tempo, a escolha de leguminosas forrageiras era limitada em condições mediterrânicas, sendo o Trevo subterrâneo (Trifolium subterraneum) e as Luzernas (Medicago spp.) as espécies mais utilizadas (Nichols, et al., 2007), podendo também encontrar-se a Tremocilha ( Lupinus luteus) e o Trevo encarnado (Trifolium incarnatum), embora em menor utilização.
Com as mudanças nos sistemas de agricultura houve a necessidade de desenvolver uma nova geração de leguminosas forrageiras, que viessem colmatar as deficiências existentes. Assim, existe hoje um maior potencial de leguminosas forrageiras melhoradas para os sistemas de agricultura mediterrânicos (Loi et al., 2005). De entre estas novas espécies desenvolvidas, incluem-se o trevo glandulifero ( Trifolium glanduliferum), o trevo vesiculoso (Trifolium vesiculosum) , o trevo balansa (Trifolium michelianum) e a senra (Biserrula pelecinus) e novas variedades de serradela vulgar (Ornithopus sativus) (Loi et al., 2005).
A data de sementeira e de corte das culturas para sideração está sempre dependente das respectivas datas de sementeira e de maturação da cultura principal. Assim, as espécies utilizadas para sideração devem ter um desenvolvimento adaptado ao período em que o solo está disponível. A temperatura e precipitação influenciam a produção de biomassa destas culturas (Sainju et al., 2005). Sementeiras tardias de culturas para sideração podem produzir entre 15 a 75% menos matéria seca quando comparadas com sementeiras mais precoces (Odhiambo e Bomke, 2001). Assim, nas nossas condições em cultivo outonal as datas de sementeira são um factor cuja importância é necessário esclarecer, já que em outras condições agro – ecológicas, o atraso na data de sementeira pode comprometer o potencial das leguminosas anuais de Inverno como culturas para sideração (Teasdale et al., 2004).
Com a introdução desta prática, aumenta-se de forma natural o teor de N no solo, o que acabará por melhorar a produtividade do solo e a qualidade ambiental, reduzindo as perdas de N por lixiviação e a emissão de N2O para a atmosfera (Sainju e Singh, 2008). Assim, é importante conhecer o efeito das culturas para sideração na mineralização do N, para desenvolver estratégias de gestão do N no solo (Schomberg et al., 2006a).
Não existem referências, na literatura, à utilização de algumas leguminosas mediterrâneas e à utilização desta técnica em Portugal, o que nos levou à elaboração de um trabalho experimental, cujos objectivos são: (1) Quantificar a produção de matéria seca das culturas forrageiras de estação fria em condições mediterrânicas; (2) Quantificar o N na biomassa dessas forrageiras anuais de estação fria; (3) Quantificar o N disponibilizado pelo cultivo de diversas leguminosas anuais de estação fria para a cultura seguinte; (4) Comparar o efeito dessas leguminosas com o azevém e com a vegetação espontânea; (5) Avaliar o efeito de duas datas de sementeira.
MATERIAL E MÉTODOS
O primeiro ano do trabalho experimental decorreu entre Setembro de 2007 e Maio de 2008 na Escola Superior Agrária, Quinta da Alagoa em Viseu (Latitude:40º40’N; Longitude 7º55’W; Altitude 450m), Portugal.
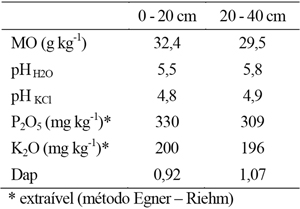
O solo onde foi instalado o ensaio apresenta reduzido declive e, portanto, baixo risco de perdas por erosão. O solo pode classificar-se como Fluvissolo dístrico (FAO, 2006). Antes do inicio da realização do ensaio efectuaram-se determinações da composição granulométrica e dos parâmetros de análise sumária, cujos resultados se apresentam nos quadros 1 e 2.
Quadro 1 - Valores médios da análise granulométrica do solo.

Quadro 2 - Valores médios da análise de rotina do solo para as profundidades 0-20cm e 20-40cm.
Os dados meteorológicos foram recolhidos na Estação Meteorológica de Viseu da Direcção Regional de Agricultura e Pescas do Centro. A precipitação média mensal e a temperatura média mensal, ocorridas durante o período que decorreu o trabalho experimental estão registadas na Figura 1.
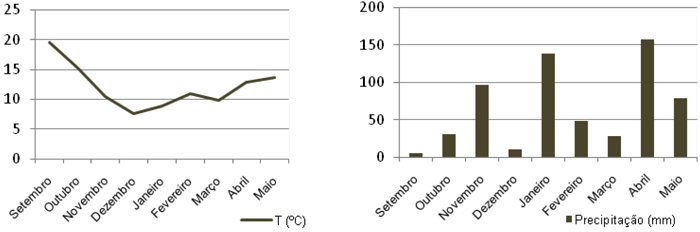
Figura 1 – Temperaturas médias mensais e Precipitação mensal em Viseu no período experimental
Os tratamentos em ensaio consistiram em nove culturas sujeitas a duas datas de sementeira. As espécies de plantas utilizadas foram o trevo balansa (Trifolium michelianum Savi) Cv. Paradana, trevo glandulífero (Trifolium glanduliferum Boiss.) Cv. Prima, trevo vesiculoso (Trifolium vesiculosum Savi) Cv. Cefalu, trevo encarnado (Trifolium incarnatum L.) Cv. Contea, serradela vulgar (Ornithopus sativus Brot.) Cv. Cadiz e tremocilha (Lupinus luteus L.) população regional como leguminosas estremes, o azevém (Lolium multiflorum Lam.) westerwoldicum diplóide Cv. Liforia e uma consociação de azevém com trevo balansa. Como tratamento testemunha foi utilizada a vegetação espontânea.
O ensaio foi delineado em talhões subdivididos ("split plot") em que os talhões principais ou grandes talhões são as datas de sementeira e os pequenos talhões (subtalhões) as diferentes espécies/modalidades de plantas. Foram efectuadas quatro repetições. A dimensão de cada subtalhão foi de 3m x 5m. É importante referir que a parcela onde se localiza o ensaio, estava já há alguns anos com vegetação espontânea.
As culturas foram semeadas em duas datas: 22 de Setembro e 27 de Outubro de 2007, pretendendo representar uma data precoce e uma data tardia, respectivamente. As densidades de sementeira utilizadas foram: para a serradela 20 kg ha-1; para o trevo balansa, trevo glandulífero e trevo vesiculoso 10 kg ha-1; para o trevo encarnado 20 kg ha-1; para o azevém 30 kg ha-1 e a tremocilha 60 sementes m-2. Na consociação utilizou-se a soma de 60% dos valores estremes, ou seja 18 kg ha-1 de azevém mais 6 kg ha-1 de trevo balansa.
Para a preparação do terreno para a sementeira foram efectuadas inicialmente uma mobilização profunda (lavoura) para destruir as infestantes e depois uma passagem com a grade de discos para preparar uma boa cama de sementeira. Aquando da lavoura foram aplicados fertilizantes de acordo com as recomendações de análises do solo realizadas para o efeito: 2000 kg ha-1 de calcário dolomítico; 250 kg ha-1 de sulfato de potássio.
O solo de cada um dos pequenos talhões de ensaio foi analisado relativamente ao N mineral imediatamente antes da instalação das culturas, aquando da 1ª data de sementeira, da 2ª data de sementeira e do corte final das culturas, antes da sideração. Para a amostragem de solos foram recolhidas cerca de 5 a 6 sub-amostras às profundidades de 0 – 20 cm e 20 – 40 cm de cada pequeno talhão, que se homogeneizaram de forma a obter uma única amostra de cada profundidade de cada pequeno talhão para posterior análise. Em laboratório, as amostras foram sujeitas a crivagem através de um crivo de malha de 2 mm para separação e determinação das proporções de terra fina e dos elementos grosseiros. Retirou-se uma amostra de cerca de 150 g que foi de imediato pesada e colocada na estufa a 105 ºC até atingir peso constante. Foi de novo pesada e posteriormente calculado o seu teor em água. Retirou-se outra amostra de cerca de 50 g que foi ultracongelada até ao momento da realização da análise do teor de N mineral no laboratório de solos da UTAD. Para extracção do N mineral utilizou-se KCl 1 M, relação 1:5 e doseamento por espectrofotometria de absorção molecular. Para análise das produções foram colhidas amostras de 0,25 m2 (0,5 x 0,5 m) em parte pré determinada de cada pequeno talhão. Foram colocadas em sacos devidamente etiquetados e levadas para o laboratório. Já no laboratório foram de imediato pesadas e registado o seu peso verde. Nas amostras relativas à vegetação espontânea e à consociação azevém x trevo balansa, fez-se a separação entre as diferentes espécies constituintes da amostra. As amostras foram então colocadas na estufa a 65 ºC até atingirem peso constante. Após este tempo foram de novo pesadas. Retirou-se uma subamostra de cerca de 150 g que foi moída (crivo de 1mm de diâmetro) para posterior quantificação de N total. Esta quantificação foi feita através de digestão por via húmida com ácido sulfúrico e o doseamento por espectrofotometria de absorção molecular em analisador de fluxo segmentado.
Após a amostragem da produção em todos os pequenos talhões do ensaio procedeu-se ao corte, destroçamento e incorporação da vegetação no solo dos respectivos tratamentos, para prosseguir com nova etapa dos objectivos experimentais. Todas as plantas de todos talhões foram cortadas no dia 07 de Maio de 2008, com auxílio de um destroçador de mato. Posteriormente, dia 08 de Maio, procedeu-se à incorporação das plantas no solo, através de uma mobilização do solo utilizando uma grade de discos.
Os resultados obtidos foram tratados por análise de variância utilizando o delineamento em talhões subdivididos ("split plot"). Para comparar as médias dos tratamentos foi utilizado o Teste de Duncan. A significância foi estabelecida para valores de probabilidade inferiores a 5%.
RESULTADOS E DISCUSSÃO
As temperaturas e a precipitação ocorridas durante o período de desenvolvimento das plantas (Setembro a Abril) influenciaram a germinação, o crescimento e a produção de biomassa das plantas, tal como referem Sainju et al. (2005). A escassa precipitação ocorrida durante o mês de Setembro e a reduzida precipitação em Outubro (Figura 1) condicionaram a emergência das plantas semeadas em Setembro, 1ª data de sementeira. Quando efectuada a análise de variância nos talhões principais em relação à produção de matéria seca não houve diferenças significativas entre as duas datas de sementeira. Esta semelhança de valores poderá ser explicada devido às condições climáticas anormais que ocorreram, pois devido à precipitação tardia as plantas semeadas em Setembro emergiram apenas poucos dias antes da sementeira de Outubro.
A ausência de diferenças na produção de matéria seca entre plantas pode ser justificada pelo facto de não haver água suficiente no solo para as plantas germinarem prontamente e se desenvolverem, o que facilitou o desenvolvimento da vegetação espontânea e que mascarou o efeito das diferentes plantas semeadas.
Apesar de as diferenças não serem significativas, pode afirmar-se que foi a vegetação espontânea a responsável pela maior produção de matéria seca na 1ª data de sementeira (5428 kg MS ha-1), seguida pela tremocilha (4910 kg MS ha-1). Na 2ª data, a espécie mais produtiva foi a serradela (7179 kg MS ha-1), tendo a vegetação espontânea registado significativamente menor produção de matéria seca (3804 kg MS ha-1). Nesta 2ª data de sementeira existem diferenças na produção de matéria seca do trevo vesiculoso e da vegetação espontânea (as produções mais baixas) quando comparados com a serradela (a produção mais elevada).
Em todos os tratamentos, com excepção do trevo vesiculoso e da vegetação espontânea, a produção de matéria seca foi superior na 2ª data de sementeira, quando comparada com a 1ª data (Figura 2). Resultados opostos foram encontrados por Odhiambo e Bomke (2001), em estudos efectuados em Vancouver, que referem as maiores produções de biomassa para as sementeiras mais precoces. Estes resultados por nós obtidos podem ser justificados pela deficiente emergência e posterior desenvolvimento das plantas devido à escassez de água no solo dos ensaios. Knight (1985) citado por Odhiambo e Bomke (2001), refere que a humidade é um factor limitante no estabelecimento do trevo encarnado. Cherr et al. (2006) referem a importância da altura em que ocorre a precipitação para o desenvolvimento de culturas de sementes pequenas.
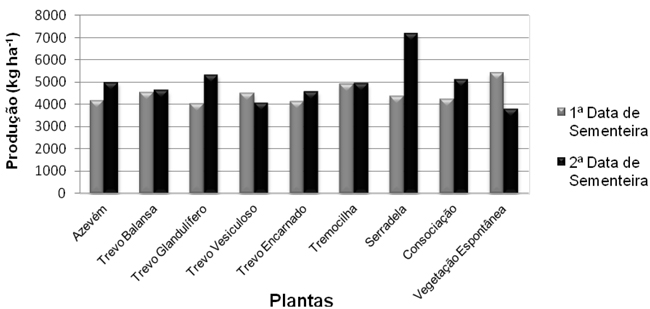
Figura 2 - Produção de matéria seca (kg ha-1) das culturas, nas duas datas de sementeira.
A maior produção de matéria seca na 1ª data de sementeira pertenceu à vegetação espontânea o que poderá ser explicado pela sua melhor adaptação/resistência às condições climáticas, nomeadamente à baixa humidade no solo para ocorrer a germinação. A produção de matéria seca na 2ª data de sementeira foi superior na consociação, quando comparada com as espécies constituintes isoladas. Resultados semelhantes foram encontrados por Sainju et al. (2005). Esta diferença é justificada por Clark et al. (1994) citado por Sainju et al. (2005), que a atribui ao facto de na consociação a densidade de sementeira ser maior, o que levará a uma redução na competição entre espécies. Em ambas as datas de sementeira o trevo encarnado obteve produções de matéria seca mais baixas quando comparadas com as outras plantas. Resultados semelhantes foram encontrados em trabalhos desenvolvidos em Vancouver (British Columbia), por Odhiambo e Bomke (2001). São apresentados por Clark (2007) valores para a produção de matéria seca do trevo encarnado entre 3923 kg ha-1 e 6164 kg ha-1. Sullivan (2003) refere valores de 4755 kg ha-1. Também para o trevo balansa, Cherr et al. (2006) apresentam valores semelhantes aos por nós obtidos: produções de matéria seca entre os 2500 kg ha-1 e os 4500 kg ha-1.
Valores ligeiramente diferentes foram apontados por Schomberg et al. (2006b), onde a produção de biomassa do trevo balansa variou entre os 1137 kg ha-1 e os 2477 kg ha-1. Em relação ao azevém, a produção de matéria seca obtida foi superior à encontrada na bibliografia. Cherr et al. (2006) apontaram valores de 1300 kg ha-1 a 2500 kg ha-1, embora Stopes et al. (1996) referidos por Cherr et al. (2006) indicassem valores de produção de matéria seca para o azevém variando entre 700 kg ha-1 e 17500 kg ha-1. Também os valores encontrados por Kuo e Jellum (2002) em Washington, são inferiores aos por nós obtidos, variando entre 1240 kg ha-1 e 1540 kg ha-1.Valores semelhantes aos encontrados na primeira data são referidos por Fowler et al. (2004), na Nova Zelândia, que não encontraram diferenças significativas na produção de matéria seca entre leguminosas, não leguminosas e consociação e apontaram valores de 4,23 t ha-1 para a gramínea, 4,85 t ha-1 para a leguminosa e 4,84 t ha-1 na consociação.
Quando efectuada a análise de variância ao teor de N nas plantas, esta evidencia a existência de diferenças muito significativas no teor de N nas plantas em função da data de sementeira e no teor de N das plantas entre as diferentes espécies/culturas. Estas diferenças entre culturas dependem da data de sementeira, pois a interacção entre as plantas e a data de sementeira foi significativa (P< 0,05). Quando comparadas as médias (utilizando o teste de Duncan) do teor de N nas plantas dentro de cada data, não foram encontradas diferenças significativas entre plantas na 1ª data de sementeira, mas foram encontradas algumas diferenças entre plantas na 2ª data de sementeira.
Quando analisados os valores das duas datas, pode verificar-se na Figura 3, que o teor de N nas plantas é muito superior em todos os tratamentos da 2ª data de sementeira quando comparado com os da 1ª data de sementeira. Na 1ª data de sementeira não existem diferenças significativas no teor de N entre as plantas. Nesta 1ª data de sementeira, foi a tremocilha que obteve um mais elevado teor de N (13,1 g kg-1), seguida pelo trevo vesiculoso (12,3 g kg-1). Os tratamentos que obtiveram os resultados mais baixos foram a consociação e a vegetação espontânea, com respectivamente 6,5 g kg-1 e 7,0 g kg-1. Para a 2ª data de sementeira houve diferenças significativas entre tratamentos. Essa diferença foi de 16,08 g kg-1 entre os valores extremos. O maior teor de N foi encontrado no trevo balansa (26,8 g kg-1), seguido pela tremocilha (23,6 g kg-1).
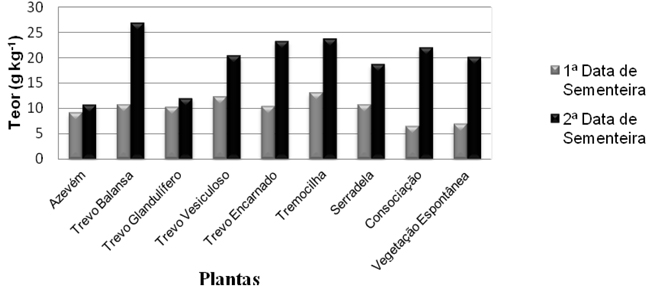
Figura 3 -Teor de N nas plantas dos tratamentos em ensaio nas duas datas de sementeira.
Os resultados mais baixos em relação ao teor de N, na 2ª data de sementeira, foram encontrados no azevém (10,8 g kg-1) e no trevo glandulífero (12,0 g kg-1). Em termos médios pode afirmar-se que o azevém e o trevo glandulífero são os mais pobres em N, enquanto que o trevo balansa e a tremocilha são os que possuem teor mais elevado. Foi no trevo balansa que se verificou a maior diferença no teor de N entre a 1ª e a 2ª data de sementeira, o que possivelmente se deve ao facto de as sementeiras da 1ª data como já referido terem sido afectadas pela falta de água após as sementeiras e a vegetação espontânea infestante se ter desenvolvido melhor. Valores semelhantes foram obtidos para o azevém por Odhiambo e Bomke (2001), que encontraram valores de 10 g kg-1 e também por Kuo e Jellum (2002), para o ano de 1998 em Washington, onde o valor foi de 10,1 g kg-1. Em todos os anos do ensaio (1994-98) estes autores encontraram os valores mais baixos de N nas não leguminosas (azevém e centeio) e os valores mais elevados nas leguminosas (ervilhaca). Valores intermédios foram encontrados pelos autores citados para a consociação (gramínea mais leguminosa). Em relação à data de sementeira, os valores encontrados por Odhiambo e Bomke (2001) não diferem muito, ao contrário do que aconteceu no presente trabalho. Esta diferença pode ser justificada pela elevada percentagem de infestantes não leguminosas, que continham os talhões da 1ª data de sementeira enquanto na 2ª data de sementeira a infestante dominante foi uma leguminosa e ainda o facto de sendo a data de corte idêntica, as plantas da primeira data de sementeira se encontrarem em estados fenológicos mais avançados.
Quando efectuada a análise de variância à quantidade de N nas culturas (parte recolhível), esta evidencia a existência de diferenças significativas na quantidade de N nas culturas em função da data de sementeira e diferenças muito significativas na quantidade de N nas culturas entre as diferentes espécies. Estas diferenças entre espécies dependem da data de sementeira, pois a interacção entre as plantas e a data de sementeira foi significativa (P<0,05). Quando comparadas as médias (utilizando o teste de Duncan) da quantidade de N das culturas foram encontradas diferenças dentro de cada data entre os tratamentos.
A quantidade de N nas culturas é superior na 2ª data de sementeira, quando comparada com a 1ª data de sementeira, como se pode observar na Figura 4.
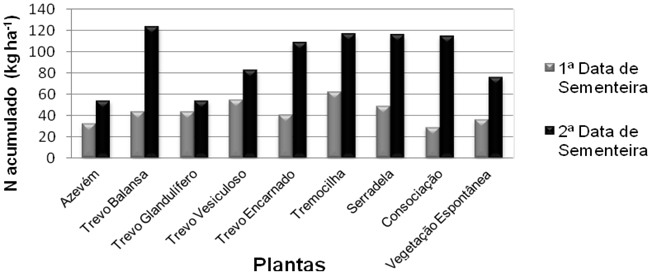
Figura 4 - Potencial de acumulação de N pelas culturas nas duas datas de sementeira.
Para a 1ª data existem diferenças significativas entre os diferentes tratamentos. Em relação à quantidade de N nas culturas, na 1ª data, foi a tremocilha que obteve o maior valor (61,8 kg ha-1), seguida do trevo vesiculoso com 55, 0 kg ha-1. As produções mais baixas, nesta data, foram encontradas na consociação (29,0 kg ha-1), seguida do azevém (32,4 kg ha-1). Na 2ª data de sementeira houve uma maior variabilidade de valores. A quantidade mais elevada de N nas culturas obteve-se no trevo balansa (123,6 kg ha-1), logo seguido da tremocilha (116,7 kg ha-1). Os valores mais baixos foram de 53,9 kg ha-1 no trevo glandulífero e de 54,0 kg ha-1 no azevém.
Em termos médios poderá afirmar-se que a tremocilha (89,29 kg ha-1) e o trevo balansa (83,74 kg ha-1) são as culturas que acumularam maior quantidade de N, sendo o azevém (43,20 kg ha-1) como gramínea estreme a cultura que acumulou menor quantidade de N.
A maior diferença de N nas culturas entre a 1ª data e a 2ª data de sementeira encontra-se na consociação que obteve apenas 29 kg ha-1 de N na 1ª data e passou para 115,08 kg ha-1 na 2ª data.
Cherr et al. (2006) citam valores de 15 a 346 kg ha-1 para o azevém, enquanto que Hanly e Gregg (2004) apontam valores de 54 a 79 kg ha-1 e Kuo e Jellum (2002) valores de 12,5 a 22,2 kg ha-1. Esta grande divergência de valores no caso do azevém está com certeza relacionada com a diferente disponibilidade de N nos solos dos diferentes ensaios e com o facto de as suas culturas serem objecto ou não de fertilização azotada. No presente ensaio não foi realizada qualquer fertilização azotada. Em todos os trabalhos os teores de N nas plantas são inferiores nas não leguminosas, tal como seria de esperar, pois todo o N que estas possuem é resultado da absorção do solo, enquanto as leguminosas conseguem também fixar o N atmosférico (Sainju et al., 2002). A inclusão de leguminosas nas culturas não leguminosas é uma forma de aumentar os valores de N dessas não leguminosas (Kuo e Jellum, 2002).
CONCLUSÕES
Este estudo mostra que as plantas utilizadas para sideração desenvolvem-se em geral com sucesso na região em estudo (Viseu, Portugal), revelando algumas das leguminosas um bom potencial de acumulação de N. De uma forma geral, as leguminosas foram mais eficientes na produção de matéria seca e na acumulação de N do que as não leguminosas. Dentro das leguminosas estudadas, foi a serradela que obteve melhores resultados para a produção de matéria seca. Na acumulação de N foi a tremocilha e o trevo balansa que retiveram a maior quantidade de N na média das duas datas de sementeira. Na sementeira mais tardia todas as plantas obtiveram maior produção de matéria seca, maior teor de N e consequentemente mais N acumulado.
Estes resultados são um bom ponto de partida para uma futura linha de investigação onde se avaliará a quantidade de N que estas plantas sideradas conseguem fornecer à cultura principal, e a forma como o disponibilizam, ou seja, se a libertação do N é feita sincronizadamente com as necessidades da cultura principal, pois as variações no teor de N da cultura principal terão origem na diferente libertação de N pelas culturas sideradas.
REFERÊNCIAS BIBLIOGRÁFICAS
Askegaard, M. e Eriksen, J. (2007) - Growth of legume and nonlegume catch crops and residual-N effects in spring barley on coarse sand. Journal of Plant Nutrition and Soil Science, 170: 773 - 780 [ Links ]
Askegaard, M. e Eriksen, J. (2008) - Residual effect and leaching of N and K in cropping systems with clover and ryegrass catch crops on coarse sand. Agriculture Ecosystems e Environment, 123: 99 – 108. [ Links ]
Bergström, L. e Kirchmann, H. (2004) - Leaching and crop uptake of nitrogen from nitrogen – 15 – labeled green manures and ammonium nitrate. Journal of Environmental Quality, 33: 1786 – 1792. [ Links ]
Cavigelli, M.A.; Teasdale, J.R. e Conklin, A.E. (2008) - Long – term agronomic performance of organic and conventional field crops in mid – atlantic region. Agronomy Journal, 100: 785 – 794. [ Links ]
Cherr, C.M.; Scholberg J.M.S. e McSorley, R. (2006) - Green manure approaches to crop production: A synthesis. Agronomy Journal, 98: 302 - 319. [ Links ]
Clark, A. 2007. Managing Cover Crops Profitably.3rd Edition. (em linha) Handbook Series Book 9. Beltsville, MD: Sustainable Agriculture Network. 246p. (Acesso em 2010.07.01) Disponivel em: < http://www.sare.org/publications/covercrops/covercrops.pdf > [ Links ]
FAO (2006) - World reference base for soil resources. Rome, Food and Agriculture Organization of the United Nations, 132 p. (World Soil Resources Reports 103). [ Links ]
Fowler, C.J.E.; Condron, L.M. e McLenaghen, R.D. (2004) - Effects of green manures on nitrogen loss and availability in an organic cropping system. New Zealand Journal of Agricultural Research, 47: 95-100. [ Links ]
Goh, K.M.; Bruce, G.E. e Sedcole, J.R. (2001) - Comparison of biological nitrogen fixation in four pairs of conventional and alternative mixed cropping farms with three rotational stages of pasture establishment in Canterbury, New Zeland. Communication Soil Science and Plant Analysis, 32 (3e4): 521 – 536. [ Links ]
Hanly, J.A. e Gregg, P.E.H. (2004) - Green-manure impacts on nitrogen availability to organic sweetcorn (Zea mays).New Zealand Journal of Crop and Horticultural Science, 32: 295 – 307. [ Links ]
Kirchmann, H.; Berström, L.; Kätterer, T.; Mattsson, L., e Gesslein, S. (2007) - Comparison of long – term organic and conventional crop – livestock systems on a previously nutrient – depleted soil in Sweden. Agronomy Journal, 99: 960 – 972. [ Links ]
Korsaeth, A. (2008) - Relations between nitrogen leaching and food productivity in organic and conventional cropping systems in long – term field study. Agriculture, Ecosystems e Environment, 127: 177 - 188. [ Links ]
Kuo, S. e Jellum, E.J. (2002) - Influence of winter cover crop and residue management on soil nitrogen availability and corn. Agronomy Journal, 94: 501 – 508. [ Links ]
Loi, A.; Howieson, J.G.; Nutt, B.J. e Carr, J. (2005) - A second generation of annual pasture legumes and their potential inclusion in Mediterranean – type farming systems. Australian Journal of Experimental Agriculture, 45, 3: 289 – 299. [ Links ]
Möller, K.; Stinner, W. e Leithold, G. (2008) - Growth, composition, biological N2 fixation and nutrient uptake of a leguminous cover crop mixture and the effect of their removal on field nitrogen balances and nitrate leaching risk. Nutrient Cycling in Agroecosystems, 82 (3): 233-249. [ Links ]
Nichols, P.G.H.; Loi, A.; Nutt, B.J.; Evans, P.M.; Craig, A.D.; Pengelly, B.C.; Dear, B.S.; Lloyd, D.L.; Revell, C.K.; Nair, R.M.; Ewing, M.A.; Howieson, J.G.; Auricht, G.A.; Howie, J.H.; Sandral, G.A.; Carr, S.J.; De Koning, C.T.; Hackney, B.F.; Crocker, G.J.; Snowball, R.; Hughes, S.J.; Hall, E.J.; Foster, K.J.; Skinner, P.W.; Barbetti, M.J. e You, M.P. (2007) - New annual and short- lived perennial pasture legumes for Australian agriculture – 15 years of revolution. Field Crops Research, 104: 10 – 23. [ Links ]
Odhiambo, J.J.O. e Bomke, A.A. (2001) - Grass and legume cover crop effects on dry matter and nitrogen accumulation. Agronomy Journal, 93: 299 – 307. [ Links ]
Olesen, J.E.; Hansen, E.M.; Askegaard, M. e Rasmussen, I.A. (2007) - The value of catch crops and organic manures for spring barley in organic arable farming. Field Crops Research, 100: 168-178. [ Links ]
Oyer, L.J. e Touchton, J.T. (1990) - Utilizing legume cropping systems to reduce N fertilizer requirements for conservation – tilled corn. Agronomy Journal, 82: 1123 – 1127. [ Links ]
Pietsch,G.; Friedel, J.K. e Freyer, B. (2007) - Lucerne management in an organic farming system under dry site conditions. Field Crops Research, 102:104 – 118. [ Links ]
Reinbott,T.M.; Conley, S.P. e Blevins, D.G. (2004) - No-tillage corn and grain sorghum response to cover crop and nitrogen fertilization. Agronomy Journal, 96: 1158 – 1163. [ Links ]
Rodrigues, M.Â.; Carneiro, J.C.; Pires, J. e Moreira, N. (2004) - Crop´s use-efficiency of nitrogen from a manure allowed for organic-farming. In European agriculture in a global context. Proceedings of the VIII ESA Congress, Copenhagen, Denmark, p. 993-994. [ Links ]
Sainju,U.M.; Sing, B.P. e Whitehead, W.F. (2002) - Long-term effects of tillage, cover crops, and nitrogen fertilization on organic carbon and nitrogen concentrations in sandy loam soils in Georgia, USA. Soil and Tillage Research, 63: 167 – 179. [ Links ]
Sainju,U.M.; Whitehead, W.F e Sing, B.P. (2005) - Biculture legume – cereal cover crops for enhanced biomass yield and carbon and nitrogen. Agronomy Journal, 97: 1403 – 1412. [ Links ]
Sainju, U.M.; Sing, B.P.; Whitehead, W.F. e Wang, S. (2007) - Accumulation and crop uptake of soil mineral nitrogen as influenced by tillage, cover crops, and nitrogen fertilization. Agronomy Journal, 99: 682 – 691. [ Links ]
Sainju,U.M. e Singh, B.P. (2008) - Nitrogen storage with cover crops and nitrogen fertilization in tilled and nontilled soils. Agronomy Journal, 100: 619 – 627. [ Links ]
Schomberg, H.H.; Endale, D.M.; Calegari, A.; Peixoto, R.; Miyazawa, M. e Cabrera, M.L. (2006a) - Influence of cover crops on potential nitrogen availability to succeeding crops in a Southern Piedmont soil. Biology and Fertility of Soils, 42: 299 – 307. [ Links ]
Schomberg, H.H.; McDaniel, R.G.; Mallard, E.; Endale, D.M.; Fisher, D.S. e Cabrera, M.L. (2006b) - Conservation tillage and cover crops influences on cotton production on a Southeastern U.S. coastal plain soil. Agronomy Journal, 98: 1247 – 1256. [ Links ]
Snapp, S.S.; Swinton, S.M.; Labarta, R.; Muthc, D.; Black, J.R.; Leep, R.; Nyiraneza, J. e O’Neil, K. (2005) - Evaluated cover crops for benefits, costs and performance within cropping system niches. Agronomy Journal, 97: 322- 332.
Sullivan, P. (2003) - Overview of cover crops and green manures (em linha). Butt, MT, ATTRA, NCAT, 16p. (Acesso em 2010.06.25) Disponivel em: < http://attra.ncat.org/attra-pub/PDF/covercrop.pdf > [ Links ]
Teasdale, J.R.; Devine, T.E.; Mosjidis, J.A.; Bellinder, R.R. e Beste, C.E. (2004) - Growth and development of hairy vetch cultivars in the North-eastern United States as influenced by planting and harvesting date. Agronomy Journal, 96: 1266- 1271. [ Links ]
Thiessen Martens, J.R.; Hoeppner, J.W. e Entz, M.H. (2001) - Legume cover crops with winter cereals in Southern Manitoba: establishment, productivity, and microclimate effects. Agronomy Journal, 93: 1086 – 1096. [ Links ]
Thorup – Kristensen, K.; Magid, J. e Jensen, L.S. (2003) - Catch crops and green manures as biological tools in nitrogen management in temperate zones. Advances in Agronomy, 79: 227 – 302. [ Links ]

