










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias v.32 n.2 Lisboa dez. 2009
A revolução verde e a biologia molecular
The green revolution and the molecular biology
Fernando Santos Henriques*
Homenagem a Norman Ernest Borlaug, Prémio Nobel da Paz, mentor e impulsionador da chamada Revolução Verde, no seu nonagésimo quinto aniversário.
Norman Borlaug faleceu no passado dia 12 de Setembro, após doença prolongada. Será recordado pelos seus familiares e pelos amigos que com ele conviveram, mas também por milhões de pessoas, homens e mulheres, em todo o mundo, pelo seu contributo decisivo para minorar os problemas da fome e da miséria nos países menos desenvolvidos, particularmente na Ásia. Até sempre, Norman.
RESUMO
No virar deste século foram identificados os genes responsáveis pela redução da estatura das variedades de trigo e de arroz que possibilitaram a chamada Revolução Verde dos anos sessenta. Mais recentemente, foi demonstrado que esta alteração da arquitectura das plantas resulta da ausência da acção das giberelinas, as hormonas responsáveis pelo alongamento dos entre-nós dos caules, mas enquanto que no trigo a redução da estatura das plantas foi o produto de mutações com ganho de função que interferem com a sequência de transdução das giberelinas, no caso do arroz resultou de uma mutação com perda de função que impede a síntese daquelas hormonas. Embora as giberelinas sejam por excelência as hormonas responsáveis pelo alongamento dos caules das plantas, é referido o caso de um mutante de milho em que a redução de altura é provocada por uma diminuição na disponibilidade de auxinas.
Estas e outras descobertas da biologia molecular vêm municiar a nova revolução requerida na agricultura para que possa satisfazer a procura mundial crescente de alimentos, sob a pressão de uma expansão contínua de culturas para produção de energia.
Palavras-chave: Auxinas, genes do nanismo, giberelinas, Revolução Verde, transgénicos.
ABSTRACT
At the turn of this century, the genes responsible for the height reduction of the wheat and rice varieties that made possible the so-called Green Revolution of the 1960s were identified. More recently, it has been shown that this change in plant architecture results from a lack of gibberellin effects, the hormones that cause the internode elongation of stems, but whereas in wheat the reduction in size was accounted for by a gain of function mutation that interfere with the signalling pathway of the gibberellins, in the rice resulted from a loss of function mutation that prevents the synthesis of those hormones. Although gibberellins are the major hormones responsible for stem elongation, it is discussed a maize mutant in which the height reduction is caused by a decrease in auxin availability.
These and other discoveries in molecular biology provide the new revolution required in agriculture with new tools in order to fulfil the global demand for increased food production, under the pressure of a continuous expansion of crops for energy production.
Key-words: Auxins, gibberellins, Green Revolution, semi-dwarfing genes, transgenics.
INTRODUÇÃO
Em meados dos anos sessenta, uma revolução invulgar tentava vingar nalguns países da Ásia, particularmente na Índia. Como outras revoluções, era feita em nome do desenvolvimento e do progresso e reclamava mudanças políticas e socioeconómicas; porém, ao contrário de outras, o seu sucesso não implicava o levantamento de massas populares, derramamento de sangue nem perda de vidas humanas. Exigia, antes, um empenhamento activo dos governos envolvidos para realizar reformas profundas nas economias nacionais de forma a poder concretizar-se o seu objectivo impar: aumentar a produção alimentar para erradicar o espectro da fome e da miséria nos países mais pobres e populosos do mundo. Não eram feitos prisioneiros, antes pelo contrário, pretendia-se libertar centenas de milhões de pessoas das grilhetas da desnutrição e do subdesenvolvimento e restituirlhes a dignidade humana. Era essencialmente uma revolução tecnológica e viria a ser conhecida por Revolução Verde 1(Gaud, 1968).
Naquela época, a maioria dos países dos continentes asiático e africano debatia-se com graves problemas de fome crónica generalizada e a ajuda internacional revelava-se não só incapaz de ultrapassar o problema de forma definitiva, como era por vezes mesmo contraproducente por baixar os preços dos produtos alimentares locais, desincentivando a produção agrícola. Tornava-se urgente encontrar vias para os próprios países beneficiários garantirem uma segurança alimentar mínima e, se possível, mesmo a auto-suficiência. Foi então que os Estados Unidos se propuseram enfrentar este enorme desafio de forma inovadora, promovendo e apoiando, técnica e financeiramente, a introdução naqueles países de novas variedades de arroz e de trigo de elevada produtividade, com forte resposta aos fertilizantes químicos, com o potencial, portanto, de reduzir ou mesmo eliminar os défices crónicos de alimentos nessas partes do mundo. A Índia tinha obtido baixíssimas produções de trigo nos anos de 1965 e 1966, devido a atraso das chuvas de monção, tendo-se visto obrigada a importar dez milhões de toneladas deste cereal dos Estados Unidos, num esforço financeiro enorme que agravou a sua balança de pagamentos e o seu endividamento externo. Era, assim, uma candidata natural para um ensaio deste novo tipo de assistência e, apesar de resistências internas, o espectro de novos episódios de fome com o risco inerente de desordens sociais levou o governo indiano a aceitar a introdução das variedades de alto rendimento.
Nos anos cinquenta, sob a orientação e o impulso do cientista e melhorador Norman Borlaug, que viria a ser distinguido com o Prémio Nobel da Paz em 1970 por este seu trabalho, investigadores da Fundação Rockefeller tinham desenvolvido no México novas variedades de trigo, de porte mais reduzido e com um potencial produtivo superior ao dobro do das variedades tradicionais. Estas variedades anãs não só concentravam os ganhos de biomassa na produção de grão (maior índice de colheita), em vez do crescimento do caule e formação de folhas adicionais, como também suportavam e respondiam melhor a elevadas doses de fertilizantes sem risco de acama, o que conjuntamente tornava possível as suas altas produtividades. Mais tarde, nas Filipinas, investigadores da mesma Fundação, juntamente com colegas da Fundação Ford, criaram igualmente variedades de arroz de palha curta, também com resposta acentuada aos fertilizantes e com produções de grão espectaculares2 (Khush, 2001). Foram estas variedades de palha curta de trigo e arroz que foram introduzidas na Índia e noutras regiões da Ásia nos anos sessenta e cujo cultivo intensivo permitiu o aumento imediato da produção alimentar, aliviando a fome de milhões de pessoas e reduzindo a necessidade de importação de cereais. Na Índia, entre 1965 e 1983 - em menos de vinte anos, portanto - a produção de trigo mais do que triplicou e o país passou de deficitário a auto-suficiente, chegando mesmo a dispor de stocks por vezes importantes. Na América Latina, onde a Revolução Verde tivera início ainda nos anos cinquenta, alguns países, como o México, por exemplo, quadruplicaram a produção de trigo entre 1950 e 1980 e passaram de importadores a exportadores de cereais3! No cômputo geral dos países em desenvolvimento, graças à Revolução Verde, a produção cerealífera aumentou 33% entre 1972 e 1982, quase o dobro do acréscimo de 18% verificado no mesmo período de tempo nos países industrializados.
Este êxito inicial da Revolução Verde criou as maiores expectativas entre os países dado-res e levou o norte-americano Lester Brown, então responsável pela assistência técnica e ajuda alimentar aos países do terceiro mundo e um dos grandes apoiantes desta experiência, num livro publicado em 1970 – Seeds of Change. The Green Revolution and Development in the 1970s – (Brown, 1970) a expressar uma grande esperança num futuro sem fome, baseado na melhoria das culturas agrícolas pelas modernas tecnologias e mudanças nas estratégias dos governos, com políticas orientadas para o crescimento do sector agrícola e aumento da qualidade de vida das suas populações. Esperança vã. Apesar dos notáveis aumentos de produção de alimentos que possibilitou, particularmente durante o período de explosão demográfica dos anos sessenta e setenta, a Revolução Verde não resolveu os problemas de fome e de miséria do mundo; infelizmente, estes problemas não só continuam actuais como se têm agravado mesmo nalguns países onde as necessidades alimentares cresceram mais do que a produção devido ao aumento populacional ou a conflitos internos. A FAO calcula que nos países em desenvolvimento mais de 950 milhões de pobres sofrem de fome crónica ou aguda (Sanchez, 2009), grande parte na Ásia – 200 milhões, pelo menos, na Índia - e, a nível mundial, cerca de um terço da população não dispõe de uma nutrição mínima adequada a levar uma vida activa saudável! A ultrapassagem desta situação requer modificações profundas em diversos domínios, principalmente na política de distribuição e acesso aos alimentos e no controlo do crescimento populacional, mas requer também, naturalmente, a continuação do aumento da produção de alimentos. Felizmente, vislumbram-se novas oportunidades de aumentos significativos da produção e produtividade agrícolas globais, através da conjugação de programas clássicos de melhoramento com a engenharia genética para a criação mais rápida de novas variedades com as características agronómicas desejadas, capazes de conseguir aquele objectivo por métodos de produção mais sustentáveis, não poluidores do solo e da água. De facto, não deve escamotear-se que a Revolução Verde, com a utilização intensiva de fertilizantes, pesticidas e herbicidas sintéticos, bem como com as copiosas irrigações que requeriam os ganhos de produção conseguidos com as variedades de alto rendimento, tiveram elevados custos para o ambiente, da degradação dos solos à poluição das águas subterrâneas, e tal deve estar presente em futuros programas de aumento da produção agrícola. Além disso, a adopção generalizada de um reduzido número de variedades de alto rendimento levou ao desaparecimento de milhares de variedades tradicionais e, consequentemente, à redução da diversidade genética destas culturas, numa perda irreversível. Finalmente, note-se ainda que dum ponto de vista socioeconómico a Revolução Verde causou um aumento das desigualdades sociais, agravou o fosso entre pobres e ricos e originou migrações maciças para as cidades (Evenson and Gollin, 2003). Em contraposição, Kesavan e Swaminathan (2006) fazem notar que se não se tivessem verificado os aumentos de produtividade das culturas de trigo e de arroz possibilitadas pela Revolução Verde, a Índia teria sido obrigada a converter cerca de 80 milhões de hectares de floresta para conseguir as produções de cereais obtidas em 2006!
As variedades de palha curta de trigo e de arroz que tornaram possível a Revolução Verde foram fruto de um trabalho árduo e longo de melhoramento genético convencional. Nos recuados anos cinquenta, o processo de produção de uma nova variedade combinando as características agronómicas desejadas era realizado pelo cruzamento de linhas parentais apropriadas, seguida de uma cuidadosa e paciente selecção da descendência, num processo geralmente demorado e de resultados imprevisíveis; por vezes, no final de muitos anos e cruzamentos, não se obtinham variedades com desempenho superior aos progenitores. De facto, durante muitas décadas, a prática de melhoramento genético era um processo largamente empírico, pois desconhecíamos não só os genes responsáveis pelas características procuradas como desconhecíamos os mecanismos de controlo da sua expressão. Ultimamente, porém, esta situação tem vindo a alterar-se progressivamente.
Identificação dos genes do nanismo do trigo e do arroz.
Passaram várias décadas até se perceber a base genética da redução do porte das plantas de trigo e de arroz que fizeram a Revolução Verde. Há cerca de dez anos, a ciência fez um progresso significativo nesta área com a identificação dos genes reguladores da altura do trigo que permitiram a produção das plantas semi-anãs - os genes Rht (de Reduced height), localizados nos cromossomas 4B (Rht-B1b) e 4D (Rht-D1b) do genoma da planta (Peng et al., 1999). Cada gene provoca uma redução semelhante na estatura da planta, mas os seus efeitos são aditivos. É importante fazer notar que os alelos Rht não afectam o crescimento dos entre-nós das espigas no período da sua expansão, de modo que o crescimento final destas é idêntico ao das plantas de estatura normal, permitindo assim as elevadas produções das plantas de altura reduzida. Estas plantas possuem mutações com ganho de função (semidominantes) naqueles genes, que se demonstrou codificarem proteínas alteradas que provocam uma ausência de resposta às hormonas promotoras do alongamento dos entre-nós, as giberelinas (GA)4; mutações em qualquer dos seus loci alteram a intensidade da resposta da planta àquela hormona e produzem graus variáveis de nanismo.
Esta foi a primeira vez que foram identificados tais genes no trigo, que se verificou serem ortólogos dos genes GAI (Gibberellic Acid Insensitive) de Arabidopsis (Koorneef et al., 1985) e dos genes D8 (Dwarf8) do milho (Fujioka et al., 1988), previamente caracterizados, para os quais também eram conhecidas mutações que produziam plantas de porte reduzido insensíveis às giberelinas5. Sabe-se que em todos os casos, as regiões amino-terminal das proteínas produzidas (gai, rht-D1 e d8), exibem alterações na sua estrutura, principalmente num domínio altamente conservado designado DELLA6, que está envolvido na sinalização das giberelinas (Fig. 1). As mutações de Rht-B1b e Rht-D1b são muito semelhantes, envolvendo ambas substituições de nucleótidos que originam codões de paragem que conduzem à formação de proteínas truncadas na região DELLA, mas que são ainda assim activas. As proteínas DELLA funcionam como repressores intracelulares (localizados principalmente no núcleo) da acção das GA, isto é, são responsáveis pela repressão da transcrição dos genes de resposta às GA na ausência da hormona. Quando esta está presente, a ligação ao seu receptor promove a interacção das proteínas DELLA com o complexo GA-receptor e induz a sua degradação, pelo que a repressão é suprimida (Fig. 2). De facto, a associação das proteínas DELLA com o complexo GA-receptor provoca a ubiquitinação daquelas proteínas e o seu subsequente reconhecimento e degradação pelo complexo proteossómico7. Os domínios DELLA são essenciais para a interacção destas proteínas com o receptor das GA na presença da hormona e, portanto, são suficientes para inactivar a função repressora destas proteínas. No caso das mutações Rht, as proteínas truncadas na região DELLA continuam a ser capazes de exercer a acção repressora dos efeitos das GA na ausência destas, mas como aquela alteração impede a sua interacção com o receptor da hormona quando esta está presente, não são degradadas, continuando a inibir a transdução do sinal GA, funcionando assim como repressores constitutivos da acção destas hormonas (V. nota 6 e figura 2).
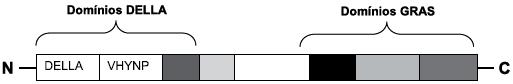
Figura 1 – Representação esquemática das proteínas DELLA, com indicação dos dois domínios da região N-terminal (DELLA e VHYNP) responsáveis pela sua acção repressora na sequência de transdução das giberelinas. Na região C-terminal localizam-se os domínios típicos da família GRAS, a que pertencem as proteínas DELLA.
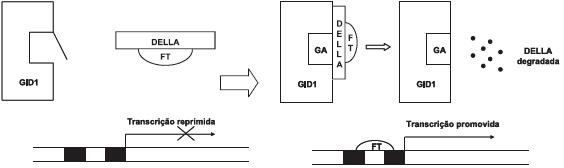
Figura 2 – Proposta do mecanismo de sinalização das giberelinas. Na ausência das GA (lado esquerdo da figura), as proteínas DELLA reprimem a expressão dos genes de resposta àquelas hormonas, possivelmente ligando-se a factores de transcrição (FT) desses genes e retendo-os em complexos inactivos; quando as GA se acumulam (lado direito da figura), as proteínas DELLA associam-se ao complexo GA-Receptor (representado na figura por GID1, receptor nuclear das GA no arroz) através de vários motivos do domínio DELLA e essa interacção vai desencadear a degradação das proteínas DELLA, eliminando a sua acção repressora e promovendo a expressão dos genes de resposta às GA.
O mecanismo pelo qual as proteínas DELLA reprimem a expressão dos genes de respostas às GA não foi ainda exactamente determinado, sendo possível que o façam por via múltiplas e, possivelmente, indirectas, por exemplo interagindo com activadores de transcrição desses genes que são assim retidos em complexos inactivos (Feng, 2008) (Fig. 2). Só em 2008 foi revelada a base estrutural do reconhecimento das GA pelo seu receptor e da interacção das DELLA com o complexo GA-receptor, em plantas de arroz (Shimada et al., 2008). No arroz, o receptor é uma proteína nuclear, GID1 (GIBBERELLIN INSENSITIVE DWARF1)8 (Ueguchi-Tanaka et al., 2005), que forma uma cavidade na sua região N-terminal, coberta por uma tampa amovível, onde se vai ligar a hormona GA, que é aí retida por interacções polares e não polares com aminoácidos de GID1. Quando a tampa se fecha, com GA no interior da bolsa, julga-se que a parte exterior da tampa, hidrofóbica, interaja com as proteínas DELLA, indo determinar a sua degradação (Fig. 2). O mecanismo que leva à ubiquitinação das proteínas DELLA e sua subsequente degradação não está esclarecido, sendo possível que envolva uma alteração conformacional no domínio GRAS da proteína, situado na região C-terminal (Fig. 2). Nalgumas plantas, as proteínas DELLA estão fosforiladas, mas o significado – se al-gum – desta fosforilação para o mecanismo de acção/degradação das proteínas permanece desconhecido (Schwechheimer et al., 2009). Mais recentemente, foi proposto que a glicosilação das proteínas DELLA é requerida para a sua acção, por estar envolvida na interacção da proteína com o receptor da hormona, mas falta suporte bioquímico para esta proposta (Schwechheimer et al., 2009).
No arroz, são também conhecidos casos de nanismo, como gid1, gid2 e slr1, por exemplo, em que os mutantes são insensíveis às GA. Contudo, a origem do nanismo das cultivares de arroz utilizadas na Revolução Verde resulta da mutação recessiva semi-dwarf1 (sd1 ou OsGA20ox-2) que causa a perda da capacidade de produção da enzima GA20oxidase (GA-20ox) que catalisa a parte final da síntese da GA20 (Monna et al., 2002; Sasaki et al., 2002). O gene, localizado no braço comprido do cromossoma 1, exibe uma ausência de 383 pares de bases que inviabiliza a síntese da enzima normal. Note-se que o genoma do arroz contém um outro gene que codifica igualmente uma GA20ox (GA20-ox1), mas este está preferencialmente expresso nas partes reprodutivas da planta, ao contrário do que sucede com sd1, que se localiza principalmente nos caules e lâmina das folhas. A mutação sd1 é a responsável pela redução da altura do arroz, enquanto o gene GA20ox-1 deve explicar a ausência de efeitos na produção de grão pelas plantas de estatura reduzida (Sasaki et al., 2002). As plântulas sd1 são, portanto, capazes de responder à aplicação exógena de GA com aumentos da sua altura, podendo mesmo atingir o tamanho das plantas normais. O gene sd1 foi originalmente introduzido de uma variedade chinesa Dee-geo-woo-gen, que foi cruzada com uma variedade de palha alta da Indonésia, Peta, em 1962. Deste cruzamento, na sequência de um programa cuidado de melhoramento, foi produzida no IRRI a nova cultivar IR8, disponibilizada aos agricultores em 1966, que permitiu que a produção de arroz duplicasse nas Filipinas no período de duas décadas! Sob condições óptimas de fertilização e irrigação, IR8 chegava a exibir uma produtividade mais de dez vezes superior às das variedades tradicionais, tendo vindo a ser utilizado não só na Ásia, mas também na África e América Latina. Refira-se, contudo, que anteriormente a Índia já criara uma nova variedade, ADT-27, que também produzia bastante melhor que as tradicionais e que estava bem adaptada às condições deste subcontinente, tendo sido esta variedade que esteve no início da Revolução Verde para a cultura do arroz neste país. IR8 foi designado o arroz maravilha e serviu de base a muitas cultivares semi-anãs de arroz indica cultivadas nas áreas tropicais e semitropicais da Ásia, mas também de arroz japonica, para cultivo nas regiões mais temperadas da República da Coreia e da Califórnia9. Contudo, IR8 não era perfeito: era atacado por algumas doenças e pragas e o grão exibia uma aparência que não era do agrado geral do consumidor. Além disso, porque continha um alto teor de amilose, endurecia depois de cozido. Por isso, logo a seguir ao seu aparecimento, começaram os esforços por criar novas cultivares sem estes inconvenientes. Surgiu primeiro IR36, uma variedade semi-anã com elevada resistência a um grande número de pragas e doenças e um ciclo de vida muito mais curto do que o de IR8 e, em 1990, foi criado o IR72 com uma produtividade superior à de IR36.
Note-se que embora a ausência dos efeitos das GA esteja na origem dos fenótipos de baixa estatura nas duas principais culturas da Revolução Verde, a base genética e a função bioquímica dos genes envolvidos são completamente distintas num e noutro caso. Lembre-se, a propósito, que o trigo tem um genoma hexaplóide, pelo que a presença de um alelo recessivo num só dos seus cromossomas dificilmente se reflectiria numa alteração visível do porte da planta, o que já é possível no arroz.
Os aumentos de produtividade conseguidos com a Revolução Verde envolveram principalmente as culturas do trigo e do arroz. Contudo, há um terceiro cereal de enorme importância agrícola e económica, o milho, sobre o qual iremos adicionar uma pequena nota. Estes três cereais, no seu conjunto, são responsáveis, de forma directa ou indirecta, por mais de 50% das calorias que ingerimos diariamente. Já se referiu acima que a mutação Dwarf8 (D8) do milho é muito semelhante à Rht-1 do trigo, sendo ambas mutações semidominantes insensíveis às GA, mas estão igualmente caracterizadas mutações sensíveis às GA, designadamente d1, d2, d3 e d5. Não há dúvida de que as giberelinas são as principais hormonas determinantes da altura da planta, mas há outras hormonas que podem conduzir a um fenótipo semelhante. Recentemente, por exemplo, foi descoberto um mecanismo pelo qual as auxinas reduzem a altura do milho (Multani et al., 2003) nos mutantes br2 (brachytic2). Esta hormona é única no sentido em que o seu transporte na planta é polar, mediado principalmente por um conjunto de proteínas de efluxo localizadas assimetricamente na membrana plásmica, designadas PIN. Foi demonstrado que o transporte basipetal (em direcção à base, descendente) das auxinas no caule do milho de br2 é reduzido e a deficiência da hormona na parte inferior do caule causa o encurtamento dos entre-nós e o consequente nanismo da planta. Plantas de milho br2 transportam uma mutação em genes responsáveis pela produção de P-glicoproteínas10, que se encontram associadas com as proteínas PIN e, possivelmente, envolvidas no transporte vectorial da hormona. Br2 é ortólogo do gene AtPGP1, cuja mutação causa igualmente alterações no transporte da auxina e redução do crescimento em Arabidopsis thaliana (Geisler et al., 2005).
Perspectivas para o melhoramento das plantas possibilitadas pela biologia molecular
A descoberta dos genes moduladores da resposta às giberelinas nas plantas de trigo e do gene responsável pelo bloqueio na síntese da hormona nas plantas de arroz, teve um enorme significado simbólico pelo papel desempenhado pelas variedades de palha curta destas duas culturas na Revolução Verde, mas é particularmente importante pelas novas e promissoras perspectivas que abriu de inserção dos genes do nanismo nos genomas de outras espécies de interesse económico. Estes genes foram já experimentalmente introduzidos noutras culturas, provocando a redução de altura e o aumento de produtividade esperados, confirmando o potencial desta nova tecnologia de modificação genética das plantas (Khush, 2001).
Até muito recentemente, as opções que se ofereciam para um necessário aumento da produção alimentar a nível global eram essencialmente duas, a saber: a expansão da área sob cultivo agrícola, feita à custa da ocupação de terrenos de floresta ou outros que desempenham funções essenciais à conservação da Natureza e, consequentemente, à preservação do frágil equilíbrio ecológico do nosso Planeta, ou o aumento de produtividade das áreas já sob cultivo agrícola, intensificando o recurso a fertilizantes, pesticidas e herbicidas, com os consequentes custos ambientais. No futuro, os ganhos de produção têm de ser conseguidos fundamentalmente por aumentos da produtividade das culturas, pois não existe grande reserva de terra por cultivar, devendo ter-se o cuidado, no entanto, de salvaguardar a qualidade do ambiente.
A biologia molecular, ao possibilitar a compreensão da base genética da produtividade das plantas e a regulação da expressão dos genes envolvidos, vem abrir uma terceira opção que não pode ser ignorada se quisermos satisfazer, de forma sustentável, a procura global crescente de alimentos. Mesmo sob condições favoráveis de crescimento, os ganhos de produtividade de muitas das principais culturas estão a aproximar-se do seu máximo e novos ganhos serão cada vez mais difíceis de conseguir. Também por isso, é essencial uma abordagem nova da problemática de como alimentar mais de 9 mil milhões de pessoas em meados do corrente século. A selecção empírica de características agronómicas de interesse, praticada durante milénios e que permitiu os impressionantes aumentos de produtividade de muitas culturas nos últimos 50 anos, deve ser agora combinada com as técnicas de engenharia genética, entre outras, que permitam a introdução de modificações específicas nas culturas, com maior eficiência e rapidez.
Para isso, é fundamental a identificação dos genes responsáveis pelas características agronómicas de interesse, sejam da redução da estatura da planta, como se discutiu acima, do aumento da produtividade fotossintética, da resistência a stresses bióticos ou abióticos até à melhoria da utilização da água – aparentemente, pelo menos, o mais complexo e difícil dos problemas em termos de biologia e o de maior relevância em termos práticos. Há alguns anos atrás, o então Secretário-Geral da ONU reclamou uma nova revolução na agricultura, que designou de Revolução Azul – more crop per drop -para realçar a grande importância do problema da escassez crescente da água para a produção agrícola.
O arroz foi o primeiro cereal cujo genoma foi completamente sequenciado, em 2005, tendo-se seguido o do sorgo (Sorghum bicolor), em 2009 – e sem dúvida que em breve conheceremos o de outros, como os do milho e trigo – mas entretanto conheceu-se o de algumas outras plantas, o que veio permitir análises genómicas comparativas que irão acelerar o conhecimento das funções e interacções dos genes. O desafio que se coloca agora é o de saber tirar partido da informação fornecida pela sequenciação dos genomas das culturas para identificar os genes responsáveis pelas características desejadas para se conseguir uma maior e melhor produção de alimentos. É expectável que muitas das características agronómicas relevantes sejam controladas por múltiplos genes, mas, surpreendentemente, tem vindo a verificar-se que tal não é necessariamente o caso e que a resistência a alguns importantes stresses é controlada por um só ou por um pequeno número de genes. Para dar apenas dois exemplos, a resistência ao encharcamento no arroz parece ser principalmente controlada por um só gene (Surridge, 2002) e a resistência à seca no sorgo é explicada, em grande parte, pela existência de cópias adicionais de poucos genes da planta (Pennisi, 2009). Reparese, portanto, que alguns importantes desafios colocados à alteração das plantas podem ser resolvidos, pelo menos em parte, manipulando de forma correcta o seu próprio genoma, sem necessidade da introdução de genes de outras espécies. Sem dúvida que a engenharia genética permite eliminar a dependência da variabilidade genética existente na espécie em causa, pela transferência de genes de espécies diferentes, até provenientes de organismos de outros reinos, criando espécies transgénicas.
O número destas espécies tem estado em constante crescimento e poderá, em breve, incluir muitas das principais culturas agrícolas, à medida que avançam as metodologias para inserção de genes. O sucesso desta tecnologia poderá ser avaliado, em parte, pela expansão da sua área de cultivo, que subiu de cerca de 3 milhões de ha em 1996 para qua-se 30 milhões em 1998 - um aumento de 10 vezes nos três primeiros anos de comercialização – e atingiu em 2008, a nível mundial, mais de 125 milhões de ha. Há uma guerra em curso entre opositores e defensores dos organismos transgénicos, mas é inquestionável que estes têm o potencial de aumentar a produtividade das culturas sem a utilização de agroquímicos poluentes, por um lado, mas também o de melhorar o valor nutritivo dos alimentos. Esta tecnologia pode oferecer novas oportunidades para se conseguir uma indispensável segurança alimentar a nível mundial que não se devem desperdiçar, sem prejuízo de se dever estar atento e alerta a eventuais perigos que transportem consigo. Muitas vezes a oposição aos transgénicos tem a ver com procedimentos de multinacionais que inviabilizam a possibilidade de estender a cultura destas plantas aos mais necessitados, os países mais pobres, ao estabelecerem preços das suas sementes que as tornam inacessíveis. Há outras preocupações válidas, como a possibilidade de cruzamento das culturas transgénicas com as indígenas e é importante que antes da generalização desta revolução biotecnológica na agricultura seja inequivocamente demonstrado que não comporta riscos inaceitáveis, nomeadamente para a saúde humana e para o ambiente. Seria desastroso trocarmos os perigos conhecidos de poluição provocados pelo uso e abuso de fertilizantes e pesticidas no passado por riscos desconhecidos e de dimensão imprevisível eventualmente causados pela utilização comercial de plantas transgénicas. Permanecerá sempre um grau de risco variável e cada caso requererá um estudo particular; na decisão final terão de ser ponderados os riscos e benefícios e optar-se pelo mal menor ou pelo bem maior, consoante as circunstâncias.
Infelizmente, a discussão das aplicações da biologia molecular à agricultura têm-se centrado, quase exclusivamente, na problemática dos transgénicos, mas é chegada a altura de olhar com outra abrangência para esta nova área do conhecimento e certamente que os resultados para a humanidade serão gratificantes.
REFERÊNCIAS BIBLIOGRÁFICAS:
Brown, Lester R. (1970) -Seeds of Change. The green revolution and development in the 1970s. Praeger Publishers (for the Overseas Development Council), New York, pp.263.
Evenson, R.E. & Gollin, D. (2003) - Assessing the impact of the Green Revolution, 1960 to 2000. Science 300: 758-762.
Feng, S.; Martinez, C.; Gusmaroli, G.; Wang, Y.; Zhou, J.; Wang, F.; Chen, L.; Yu, L.; Iglesias-Pedraz, J.; Kirchner, S.; Schäfer, E.; Fu, X.; Fan, L. & Deng, X. (2008) - Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature 451: 475-479.
Fujioka, S.; Yamane, H.; Spray, C.; Katsumi, M.; Phinney, B.; Gaskin, P.; MacMillan, J. & Takahashi, N. (1988) - The dominant nongibberellin-responding dwarf mutant (D8) of maize accumulates native gibberellins. Proc. Natl. Acad. Sci. 85: 9031-9035.
Gaud, William S. (1968) - The green revolution: accomplishments and apprehensions. Discurso perante a Society for International Development, Washington, DC.
Geisler, M.; Blakeslee, J.; Bouchard, R.; Lee, R.; Vincenzetti, V.; Bandyopadhyay, A.; Titapiwatanakun, B.; Peer, A.; Bailly, A.; Richards, E.; Ejendel, K.; Smith, P.; Baroux, C.; Grossniklaus, U.; Müller, A.; Hrycyna, A.; Dudler, R.; Murphy, S. & Martinoia, E. (2005) - Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1. PlantJ. 44: 179-194.
Kesavan, P.C. & Swaminathan, M. S. (2006) -From Green revolution to Evergreen revolution: pathways and terminologies. Cur. Sci. 90: 145-146.
Khush, Gurdev (2001) - Green revolution: the way forward. Nat. Rev. Gen. 2: 615-622. [ Links ]
Koorneef, M.; Elgersma, A.; Hanhart, C.J.; van LOenen- Martinet, E.P.; van Rij. L. & Zeevaart, J.A.D. (1985) - A gibberellin insensitive mutant of Arabidopsis thaliana. Physiol. Plant. 65: 33-39.
Monna, L.; Kiytazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S. & Minobe, Y. (2002) -Positional cloning of rice demidwarfing gene, sd-1: Rice Green Revolution encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 9: 11-17.
Multani, D.S.; Briggs, S.P.; Chamberlin, M.A.; Blakeslee, J.J.; Murphy, A.S.; Johal, G. S. (2003). Loss of an MDR Transporter in Compact Stalks of Maize br2 and Sorghum dw3 Mutants. Science 302: 234-237.
Peng, J.; Richards, D.; Hartley, N.; Murphy, G.; Devos, K.; Flintham, J.; Beales, J.; Fish, L.; Worland, A.; Pelica, F.; Sudhakar, D.; Christou, P.; Snape, J.; Gale, M. & Harberd, N. (1999) - Green Revolution genes encode mutant gibberellin response modulators. Nature 400: 257-261.
Pennisi, E. (2009) - How sorghum withstands heat and drought. Science 323: 573-574.
Sanchez, P. (2009) - A smarter way to combat hunger. Nature 458: 148.
Sasaki, A., Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.; Kitano, H. & Matsuoba, M. (2002) -A mutant gibberellin-synthesis gene in rice. 2002. Nature 416: 701-702.
Schwechheimer, Claus & Willige, Björn C. (2009) -Shedding light on gibberellic acid signalling. Cur. Op. Plant Biol. 12: 57-62.
Shimada, A.; Ueguchi-Tanaka, M.; Nakatsu, T.; Nakajima, M.; Naoe, Y.; Ohmiya, H. & Matsuoka, M. (2008) - Structural basis for gibberellin recognition by its receptor GID1. Nature 456: 520-523.
Surridge, C. (2002) - The rice squad. Nature 416: 576-578.
Ueguchi-Tanaka, M.; Ashikari, M.; Nakajima, M.; Itoh, H.; Katoh, E.; Kobayashi, M.; Chow, T.; Hsing, Y.; Kitano, H.; Yamaguchi, I & Matsuoka, M. (2005) - Gibberellin insensitive dwarf1 encodes a soluble receptor for gibberellin. Nature 437: 693-698.
1 A expressão Revolução Verde (Green Revolution) foi utilizada pela primeira vez em 1968, por William Gaud, então Director da USAID, numa comunicação à Sociedade Internacional para o Desenvolvimento, em Washington, DC.
2 Em 1960, as Fundações Rockefeller e Ford criaram, conjuntamente, o International Rice Research Institute (IRRI), em Los Banos, nas Filipinas, com o objectivo de desenvolver novas variedades de arroz que pudessem aumentar a produção destecereal em países como a Índia e o sudeste asiático; muitas dessas variedades, porém, viriam também aser utilizadas na África e América Latina.
3 Em 1943, o México importava metade do trigo que consumia; em 1956 tornou-se auto-suficiente e, em 1964, exportou meio milhão de toneladas deste cereal! As variedades de trigo de alto rendimento utilizadas para se conseguirem estes aumentos de produção foram desenvolvidas no Centro Internacional de Mejoramiento de Maiz e Trigo (CIMMYT), sob a orientação de N. Borlaug.
4 As GA formam uma enorme família de diterpenóides tetracíclicos, de que se conhecem mais de 130 membros, mas apenas algumas são biologicamente activas, designadamente as GA1, GA3 e GA4.
5 As plantas mutantes que exibem uma redução da altura podem ser agrupados em duas classes, (1) mutantes sensíveis às GA, em que o nanismo resulta de uma interferência na síntese destas hormonas, sendo possível restaurar o tamanho normal por aplicação exógena das mesmas; (2) mutantes insensíveis às GA, em que a aplicação da hormona não causa nenhum ou apenas um reduzido efeito no crescimento da planta. As plantas de trigo com mutações de Rht pertencem a esta última categoria e acumulam mesmo, muitas vezes, altos níveis de giberelinas bioactivas. Rht-1 e Rht-2 foram originalmente introduzidos a partir de uma variedade de trigo japonesa semi-anã, Daruma, que foi cruzada com variedades norte-americanas de alto rendimento para produzir a cultivar Norin 10. Norin 10 foi depois utilizada em programas de melhoramento e um dos seus cruzamentos, Norin 10-Brevor 14 (NB14), foi enviado para Norman Borlaug, no CIMMYT, onde foi utilizado para cruzamentos com variedades adaptadas às regiões tropicais e subtropicais para originar os trigos utilizados na Revolução Verde (Hedden), sendo igualmente a origem do nanismo de muitas variedades de trigo comerciais.
6 As proteínas que partilham este domínio altamente conservado na região amino-terminal são designadas DELLA (das iniciais dos 5 primeiros aminoácidos do domínio) e constituem um subgrupo da família de proteínas GRAS (de GAI, RGA e SCR, os nomes dos três primeiros membros da família identificados), única das plantas, que funciona como importante reguladora de diversos processos celulares, designadamente da sinalização de hormonas.
7 O proteossoma é um complexo proteolítico que degrada proteínas marcadas pela ubiquitina. Os substratos marcados são reconhecidos por um componente do complexo (RP) e depois transferidos para uma câmara central (CP) onde são degradados em pequenos péptidos, num processo dependente do ATP.
8 Em Arabidopsis, por exemplo, foram já identificados três genes ortólogos de GID1 – AtGID1a, AtGID1b e AtGID1c – que desempenham funções redundantes na recepção das giberelinas.
9 O arroz (Oryza sativa L.) é normalmente dividido em dois subgrupos (por vezes, também referidos como sub-espécies), indica e japonica, comercialmente conhecidos como arroz do tipo agulha e carolino, respectivamente.
10 P-glicoproteínas são proteínas transmembranares com actividade transportadora da família ABC (ATP-Binding Cassette), que nos animais tem sido demonstrado estarem envolvidas no efluxo de drogas utilizadas no combate a tumores e, portanto, responsáveis por resistência ao tratamentodesta doença. É interessante que nas plantas a presença destes transportadores aumenta igualmente a resistência a herbicidas e outros compostos químicos tóxicos, sugerindo uma função idêntica à já conhecida nos animais.
* Secção de Biologia Vegetal, Departamento de Ciências da Vida, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa, Quinta da Torre, 2829-516 Caparica. E-mail: ffh@fct.unl.pt
Recepção/Reception: 2009.07.08
Aceitação/Acception: 2009.08.13

