










Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Ciências Agrárias
Print version ISSN 0871-018X
Rev. de Ciências Agrárias vol.32 no.2 Lisboa Dec. 2009
Concentrações e fluxos de nutrientes no gotejo e escorrimento do tronco de Fraxinus angustifolia Vahl
Nutrient concentrations and fluxes in the throughfall and stemflow of Fraxinus angustifolia Vahl
Ermelinda Pereira1, Manuel Madeira2, Francisco Gonçalves de Abreu2
RESUMO
Avaliou-se a concentração e os fluxos de nutrientes no gotejo e no escorrimento do tronco de freixo (Fraxinus angustifolia Vahl), num lameiro do Nordeste de Portugal. A precipitação bruta, a precipitação sob coberto (throughfall) e o escorrimento do tronco (stemflow) foram objecto de monitorização e amostras foram colhidas periodicamente para determinação do pH, dos teores de carbono (COD) e azoto (NOD) orgânicos dissolvidos e de nutrientes. Os fluxos anuais de COD, NOD, Cl, K, Na, Ca, Mg e P na precipitação bruta foram, respectivamente, 3.93, 0.92, 0.20, 0.06, 1.81, 2.40, 1.11, 1.16, 0.29 e 0.06 g m-2, enquanto no gotejo foram de 4,42, 0,20, 1,82, 1,82, 0,31, 0,54, 0,62, 0.20 g m-2; o fluxo de N-NH4+ foi nulo e o de N-NO-3 foi negativo (-0,03 g m-2). A expressão dos fluxos devido ao escorrimento do tronco variará com a área influenciada por esta solução. A restituição de nutrientes ao solo pelo gotejo e pelo escorrimento do tronco poderá contribuir para diferenciar as características do solo sob a copa das árvores.
Palavras-chave: Freixo, gotejo, nutrientes, precipitação bruta, solo.
ABSTRACT
The effect of the ash-tree (Fraxinus angustifolia Vahl) crown on characteristics of throughfall and stemflow solutions was assessed in wet pastures located in north eastern Portugal. The amounts de gross rainfall, throughfall and stemflow were measured continuously and samples were periodically collected to determine pH, dissolved organic C (DOC) and N (DON), and nutrients. The annual fluxes of DOC, DON, Cl, K, Na, Ca, Mg and P in gross rainfall were, respectively, 3.93, 0.92, 0.20, 0.06, 1.81, 2.40, 1.11, 1.16, 0.29 and 0.06 g m-2, while those regarding throughfall were 4.42, 0.20, 1.82, 1.82, 0.31, 0.54, 0.62 and 0.20 g m-2; the flux of N-NH4+ was nil and that of N-NO-3 was negative (-0.03 g m-2). The fluxes in relation with stemflow were small if expressed on a tree crown projection area basis, but can be much greater if the actual area affected is considered. The amounts and distribution of nutrients turned to the soil by throughfall and stemflow may lead to soil chemical characteristics differentiation beneath tree crowns.
Key-words: Ash-tree, gross rainfall, throughfall, nutrients, soil.
INTRODUÇÃO
Os lameiros com árvores espaçadas constituem um sistema característico das paisagens do Nordeste de Portugal. Esse sistema, como o montado (Nunes et al., 2003), caracterizase na generalidade por uma estrutura espacial heterogénea, permitindo diferenciar áreas claramente influenciadas pelas copas da árvores, de outras onde os múltiplos efeitos destas, em caso de existir, se fazem notar de forma débil e indirecta. Estudos em sistemas semelhantes relatam que as árvores espaçadas podem levar à ocorrência de ilhas de fertilidade devido ao acréscimo de inputs de resíduos orgânicos e de nutrientes que transferem para o solo via queda de folhas, senescência das raízes, gotejo e escorrimento ao longo do tronco. Segundo Gersper & Holowaychuk (1970) e Seiler & Matzner (1995), o escorrimento do tronco e o gotejo podem determinar gradientes de humidade e de nutrientes a partir do tronco das árvores e assumir um papel importante na diferenciação espacial das características físicas e químicas do solo.
O papel da copa das árvores na modificação das características químicas da precipitação tem sido documentado em vários estudos referentes a diversas espécies florestais (Parker, 1983; Gauquelin et al., 1992; Seiler & Matzner, 1995; Rodá et al., 2002; Nunes et al., 2003). O fluxo de nutrientes através do gotejo é frequentemente atribuído à precipitação, deposição seca e lixiviação dos solutos intracelulares da folhagem. No entanto, alguns nutrientes não mostram esse enriquecimento porque são absorvidos pela folhagem e líquenes (Moreno et al., 2001). A discriminação dos processos que ocorrem na copa torna-se importante, uma vez que a deposição húmida e seca corresponde frequentemente a inputs externos para o sistema, enquanto que a lixiviação representa a recirculação de nutrientes dentro do sistema (Rodrigo et al., 2003).
Em Portugal foram efectuados alguns estudos em sistemas florestais ou agroflorestais, em que se analisou o contributo da precipitação e do gotejo no ciclo de nutrientes, destacando-se os de Portela & Pires (1995) num souto de Trás-os-Montes, de Cortez (1996) e de Bispo (2002) em eucaliptais na área de Pegões; de Cortez (1996) num pinhal na área da Companhia das Lezírias, bem como de Nunes et al. (2003) em montados de Quercus rotundifolia Lam. no Sul de Portugal (região de Évora). Nestes sistemas também foram estudados os efeitos do escorrimento do tronco das árvores na variação espacial das características do solo (Cortez et al., 2004). Porém, nenhuma informação existe no respeitante ao freixo, mormente quando se pretende analisar, em detalhe, o efeito da presença desta espécie nos lameiros do Nordeste de Portugal.
Neste contexto, foi desenvolvido um estudo num lameiro localizado no Nordeste de Portugal (região de Bragança) para avaliar o efeito das copas de freixo (i) na intercepção da precipitação bruta e nas características químicas do gotejo e do escorrimento do tronco e (ii) no fluxo de nutrientes para o solo e a sua possível influência na diferenciação das características do mesmo. A informação respeitante ao efeito na intercepção da precipitação bruta foi relatada em publicação anterior (Pereira et al., 2007).
MATERIAL E MÉTODOS
Área experimental
O estudo experimental foi efectuado num lameiro localizado junto à aldeia de Alfaião (41º 4655N; 6º 2451W; 510 m de altitude), pertencente ao concelho de Bragança, no Nordeste de Portugal. Este lameiro tem uma área aproximada de 3 hectares e situa-se num pequeno vale assente em formações silúricas do complexo vulcano silicioso (Pereira et al., 2000), enquadrado numa paisagem de relevo bastante ondulado. Os solos da área do lameiro correspondem a Fluvissolos êutricos (FAO/UNESCO, 1988). O clima da região é do tipo mediterrânico, caracterizado por elevada secura e temperatura no período estival, enquanto no Inverno é chuvoso e muito frio. Em Bragança, que dista cerca de 12 km do local de estudo, a precipitação média anual é de 741,1 mm (85,9 % no período de Outubro a Maio) e a temperatura média anual de 11,9ºC (INMG, 1991). Na Figura 1 apresenta-se os valores mensais da precipitação bruta e a média mensal da temperatura do ar ao longo do período de estudo, constatando-se que durante parte deste (Novembro de 2000 a Março de 2001) a precipitação foi muito superior à normal climatológica.
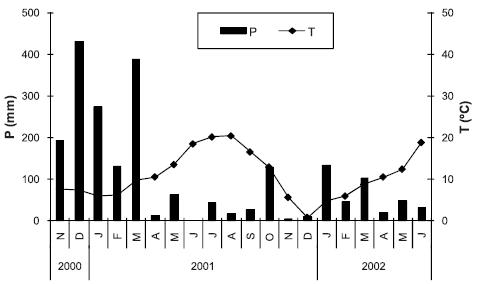
Figura 1 – Valores mensais da precipitação (P) e média mensal da temperatura (T) do ar durante o período de estudo.
Amostragens
A quantificação da precipitação bruta e da precipitação sob coberto foi realizada por intermédio de udómetros de pratos basculantes, modelo ARG 100 da Environmental Measurements, fixos a um estrado de madeira e com a boca a 60 cm do solo. Sob o coberto de uma árvore de freixo, que apresentava uma altura total de 17,0 m, uma altura da copa de 14,7 m, um diâmetro do tronco à altura do peito de 0,4 m e uma área de projecção vertical da copa de 104,9 m2, instalaram-se 4 udómetros a cerca de 1,50 m do tronco, distribuídos segundo os eixos cardeais principais (N, S, E, W); instalou-se um udómetro fora da influência da copa da árvore para medir a precipitação bruta. Os udómetros foram ligados a um sistema de registo e armazenamento de dados, modelo CR10 da marca Campbell Scientific, programado para registar a precipitação acumulada horária e diária. Ao lado de cada udómetro colocou-se um garrafão com capacidade de 5 L, ancorado ao solo através de duas estacas de ferro, que suportava no topo um funil de polietileno com 19 cm de diâmetro, para recolha da precipitação. O funil foi rodeado por uma rede pontiaguda em torno da sua extremidade mais larga, de forma a evitar que as aves nele se empoleirassem, e no seu interior foi colocado uma rede de malha de 1mm para evitar a entrada de resíduos. Estes garrafões foram envoltos por um saco de plástico preto de forma a evitar o desenvolvimento de algas. No final do período de cada amostragem, normalmente de duas semanas, quantificaram-se as precipitações acumuladas e recolheram-se amostras de água para processamento analítico. Após a quantificação e colheita de amostras, em cada data de amostragem, os udómetros foram lavados com água destilada, após o que se aplicou 1 mL de clorofórmio para impedir o desenvolvimento de algas.
A colheita do escorrimento do tronco (ET) foi efectuada através de uma caleira helicoidal de borracha colocada à volta do tronco da árvore de forma que a água escorrida ao longo do tronco fosse conduzida para um recipiente, com uma capacidade de 50 L. O volume de água medido via ET foi referido à área da projecção vertical da copa da árvore. A estimativa da quantidade de ET que atingiu o solo foi também calculada dividindo o volume obtido durante o período de estudo, pela suposta área de contribuição, considerando um gradiente de distância (10, 25, 50, 100 e 150 cm) em relação ao tronco da árvore.
Para a comparação entre a concentração de nutrientes (e valores de pH) na precipitação bruta, no gotejo e no escorrimento do tronco considerou-se, no caso do gotejo, em cada data de amostragem, a média dos quatro locais de amostragem (N, S, E, W) sob a copa da árvore.
Métodos analíticos
As soluções correspondentes à PB, GO e ET, após a sua chegada ao laboratório, foram filtradas sob um sistema de vácuo, através de uma membrana de nitrato de celulose com uma porosidade de 0,45 µm de diâmetro e em seguida armazenadas, a uma temperatura de –18ºC, em câmara frigorífica até à realização das análises.
Os teores de Ca e Mg nas soluções, após diluição das soluções com cloreto de estrôncio, foram determinados por espectrofotometria de absorção atómica. As determinações do K e Na foram efectuadas directamente nas soluções por fotometria de emissão de chama. Os teores de P e de azoto orgânico dissolvido (NOD) foram medidos num autoanalizador fluxo contínuo e o teor de carbono orgânico dissolvido (COD) num autoanalizador de carbono (DOC). Os teores de N-NO3-e de NNH4+ foram determinados por espectrofotometria de absorção molecular, em autoanalizador automático de fluxo segmentado (Houba et al., 1994). A determinação do pH efectuouse directamente nas soluções utilizando um potenciómetro da marca MetrohmHM.
Cálculos e métodos estatísticos
O cálculo dos fluxos de nutrientes devido à precipitação bruta, ao gotejo e ao escorrimento do tronco foi determinado através do somatório do produto do volume de cada solução pela concentração de nutrientes, correspondentes aos vários eventos de precipitação considerados. A comparação estatística entre a concentração de nutrientes na precipitação bruta, na precipitação sob coberto e escorrimento do tronco efectuou-se por intermédio de testes não paramétricos H de Kruskal-Wallis, dado não ter sido possível normalizar e homogeneizar as variâncias das amostras. As diferenças foram consideradas significativas para p< 0,05.
RESULTADOS E DISCUSSÃO
Características das soluções
Carbono e azoto orgânicos dissolvidos
A concentração média de carbono orgânico dissolvido (COD) na precipitação bruta (PB, 4,5 mg L-1) foi baixa (Quadro 1) e inferior à registada por Moreno et al. (2001) em quatro sítios, considerados não poluídos, na Província de Salamanca (5,9 a 7,1 mg L-1), em Espanha, sugerindo que na área do presente estudo não existirão fontes relevantes de emissão de carbono. As concentrações de COD no gotejo e no escorrimento ao longo do tronco (18,2 e 58,3 mg , respectivamente) foram significativamente mais elevadas do que na PB. Estes acréscimos seguem o padrão observado no estudo de Moreno et al. (2001) em Quercus rotundifolia, atribuindose a processos de lavagem e lixiviação de constituintes orgânicos dos tecidos vegetais.
Quadro 1 – Valores médios de pH e das concentrações médias ponderadas de C (COD) e N (NOD) orgânicos dissolvidos e de nutrientes na precipitação bruta (PB), no gotejo (GO) e no escorrimento ao longo do tronco (ET) de freixo, entre Dezembro de 2000 e Junho de 2002.
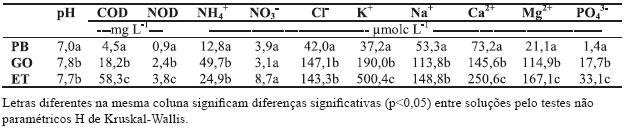
A concentração de azoto orgânico dissolvido (NOD) no gotejo (2,4 mg L-1) também foi significativamente maior do que na PB (0,9 mg L-1). Porém, o enriquecimento observado para o NOD foi menos acentuado do que o estimado para o COD, dado que a razão COD/NOD aumentou da PB (5,0) para o GO (7,6). A razão COD/NOD na solução de ET também foi muito mais elevada (15,3) do que na PB e GO, sugerindo uma maior lavagem e/ou lixiviação de C relativamente ao NOD.
Concentração de espécies iónicas
A concentração média ponderada de NH+4 (12,8 µmolc L-1 na PB foi significativamente menor do que no GO e no ET (respectivamente, 49,7 e 24,9 µmolc L-1; Quadro 1), o que se atribui à deposição atmosférica (húmida e seca), lavagem e lixiviação da copa das árvores (Moreno et al., 2001). Os acréscimos observados corroboram os relatados por Bispo (2002) e Nunes (2004) em estudos realizados no Sul de Portugal, em E. globulus e Q. rotundifolia, respectivamente. Contrastam, porém, com os observados por Moreno et al. (2001), em Q. pyrenaica, e por Rodrigo et al. (2003) em Q. ilex, que observaram uma diminuição da concentração de NH+4 no GO e que atribuíram à absorção deste catião pela folhagem e, possivelmente, à retenção pela flora epífita. As concentrações de NH+4 no gotejo correlacionaram-se positivamente com as de NOD (r = 0,75; p<0,001), reflectindo um padrão de variação comum entre elas.
A concentração de NO-3 na PB (3,9 µmolc L-1) foi ainda mais baixa do que as relatadas em estudos efectuados em áreas distantes de fontes antropogénicas, por Moreno et al. (2001) na Serra de Gata (entre 7,2 a 9,5 µmolc L-1), por Bispo (2002) na área de Pegões (29,0 µmolc L-1) e por Rodrigo et al. (2003) próximo de Barcelona (7,9 µmolc L-1). Ao contrário do observado para o NH+4, o NO-3 apresentou um ligeiro decréscimo de concentração da PB para o GO, o que deverá estar em correspondência com a possível absorção deste anião pela folhagem de freixo. Esta tendência está em acordo com o facto da concentração de NO3 - ter aumentado da solução de GO para a de ET, embora as diferenças não tenham sido significativas (p>0,05). O acréscimo de concentração observado no ET em relação à PB, devido à lixiviação e lavagem do tronco, foi também observado, no nosso País, em plantações de E. globulus (Cortez, 1996) e em montados de Q. rotundifolia (Nunes, 2004), e na Europa Central em florestas de F. sylvatica (Meiwes et al., 1998).
A concentração de Cl-observada na PB (42,0 µmolc L-1) foi cerca de um terço da determinada por Bispo (2002) a 30 km da costa Atlântica (135 µmolc L-1), atribuindo-se a diferença à grande distância da área do presente estudo à costa Atlântica. De facto, as elevadas concentrações de Cl-na atmosfera estão normalmente associadas à proximidade do mar, diminuindo à medida que a distância à costa aumenta, e à direcção dominante do ventos, os quais são responsáveis pela colocação em suspensão na atmosfera deste aerossol (Attiwill & Leeper, 1987; Aber & Melillo, 2001). As concentrações de Cl-aumentaram significativamente na solução do GO e ET em relação à PB. Este aumento está em linha com resultados obtidos na península Ibérica por Moreno et al. (2001) em Q. pyrenaica, por Bispo (2002) em E. globulus e por Rodrigo et al. (2003) em Q. ilex, devendose à deposição seca no coberto vegetal, dado considerar-se que o Cl-não é absorvido ou lixiviado pelos tecidos vegetais (Attiwill & Leeper, 1987).
As concentrações dos catiões básicos (Na+, K+, Mg2+ e Ca2+) também foram significativamente maiores no GO (113,8, 190,0, 114,9 e 145,6 µmolc , respectivamente) do que na PB (53,3, 37,2, 21,1 e 73,2 µmolc L-1, respectivamente). No ET, á excepção do Na+, as concentrações destes catiões foram significativamente mais elevadas do que no GO (Quadro 1). Os maiores acréscimos no gotejo corresponderam ao Mg2+ e ao K+, enquanto no ET o acréscimo de concentração de K+ foi muito maior do que o dos outros catiões básicos (Quadro 2). Este padrão reflecte o facto do K+ apresentar uma grande mobilidade nos tecidos vegetais e, por isso, ser fortemente lixiviado dos mesmos (Marschner, 1995). Os acréscimos de concentração dos catiões básicos no GO e ET seguem, globalmente, a tendência observada por Portela & Pires (1995) em C. sativa, por Nunes (2004) em Q. rotundifolia e por Moreno et al. (2001) em Q. pyrenaica, estando associados à lixiviação e lavagem das copas.
Quadro 2 – Razões entre as concentrações dos iões no gotejo (GO) e na precipitação bruta (PB) e entre as observadas no escorrimento ao longo do tronco (ET) e na preciptação bruta (PB).

A concentração de PO43-na PB (3,8 µmolc L-1) afastou-se bastante da observada por Portela & Pires (1995) em soutos (1,29 µmolc L-1) da região de Trás-os-Montes (Carrazedo de Montenegro), e por Rodrigo et al. (2003) em dois locais no NE de Espanha (1,03 e 1,25 µmolc As elevadas concentrações de PO4 observadas ao longo do período de estudo poderão estar relacionadas com fontes de emissão locais de partículas do solo provenientes, sobretudo, da actividade agrícola, bem como da actividade da avifauna (Ludwig, 2001). Comparativamente aos outros iões estudados, a concentração de PO43-foi a que mais aumentou pela passagem da água da chuva pela copa e tronco de freixo, sendo cerca de 13 vezes mais elevada no GO e 24 vezes no ET (Quadro 2). O acréscimo de concentração de PO43-no ET em relação ao GO não corrobora o padrão observado por Portela & Pires (1995), por Cortez et al. (2004) e por Rodrigo et al. (2003) em estudos com outras espécies florestais. Verificouse, ainda, que as concentrações de PO43-no GO e ET se correlacionaram positivamente com os teores de (r = 0,67, p<0,01; r=062, p<0,01), reflectindo semelhança de comportamento na planta. Tal facto deverá estar em correspondência com o elevado teor de P nas folhas verdes e folhas senescentes do freixo (Pereira, 2004), em que a razão N/P é inferior a 10, corroborando também as considerações de Marschner (1995) acerca da mobilidade e lixiviação de P dos tecidos vegetais. Aliás, a correlação positiva entre as concentrações PO43-e de NOD no gotejo (r = 0,79; p<0,001), reflectindo um padrão de variação comum, também atestam a hipótese de elevada mobilidade do primeiro na folhagem do freixo. Assim, as diferenças entre o freixo e as outras espécies, para além da concentração e formas de P na folhagem, poderão dever-se às características morfológicas de cada uma, nomeadamente a inclinação e densidade dos ramos, ou ainda características locais que determinam a maior ou menor deposição das partículas do solo. Aliás, segundo Levia & Herwitz (2002) o ângulo de inclinação dos ramos determina a maior ou menor concentração de nutrientes, que é atribuído ao tempo de contacto da água da chuva com a superfície dos ramos.
Valores de pH
O valor médio de pH da PB (7,0) foi bastante mais elevado do que os observados em Portugal, em áreas também consideradas de reduzida influência antropogénica. Com efeito, esse valor é, de facto, superior ao determinado por Cortez (1996) e Bispo (2002) na área de Pegões (5,35-5,73) e Cortez (1996) na Companhia das Lezírias (5,36), na área de Porto Alto; também foi superior aos observados por Moreno et al. (2001), na Província de Salamanca (5,35 a 5,58), e por Rodrigo et al. (2003) no NE de Espanha (6,58). É também mais elevado do que o relatado por Nunes (2004) na região de Évora (6,16). Os elevados valores de pH obtidos no presente estudo poderão ser justificados pelo baixo impacto das acções antropogénicas na região no que se refere à actividade industrial. Com efeito, as concentrações de NO3 e NH4+ -iões responsáveis pela acidez da precipitação e presentes em concentrações elevadas em áreas sob forte influência antropogénica (Casartelli et al., 2008) - foram reduzidas e as concentrações de catiões básicos elevadas (Quadro 1); aliás, o valor de pH referido também sugere que as concentrações de SO42-devem ser diminutas.
Os valores médios de pH na solução do gotejo (7,8) aumentaram de forma significativa (p<0,05) em relação à PB, correspondendo a uma solução claramente alcalina; tais valores seguindo a tendência observada por Cortez (1996) Moreno et al. (2001) e Nunes (2004) em várias espécies florestais. A alteração do pH sob o coberto depende da modificação do balanço entre espécies aniónicas e catiónicas, devido à influência da deposição seca e da absorção ou lixiviação das mesmas pelas copas, conforme os casos (Bispo (2002; Casartelli et al. 2008). No caso do presente estudo poderá estar relacionado com a influência da lavagem e lixiviação dos catiões básicos dos tecidos vegetais.
No presente estudo não foi observada a diminuição dos valores de pH na solução do ET em relação à PB, como concluem os estudos de Cortez (1996) em E. globulus e P. pinaster e de Moreno et al. (2001) em Q. rotundifolia. Pelo contrário, observouse um aumento (p<0,05) em cerca de 7 décimas em relação à PB e um aumento não significativo em relação ao gotejo (p>0,05) (Quadro 1). Esta tendência corrobora aquela observada por Nunes (2004), em Q. rotundifolia, e por Rodrigo et al. (2003) em Q. ilex.
Fluxos de nutrientes
Os fluxos anuais de alguns elementos provenientes da atmosfera (deposição húmida e seca) diferem grandemente dos registados noutras regiões do País. Por exemplo, os fluxos anuais de Na e Cl (1,11 e 1,81 g m-2 , respectivamente) via precipitação bruta são menores do que os observados por Cortez (1996) e Bispo (2002) na área de Pegões, o que reflecte a maior distância da área do presente estudo ao oceano. Porém, os fluxos anuais de Ca, Mg e K (1,16, 0,29 e 2,40 g m-2, respectivamente), no presente estudo (Quadro 3), foram maiores do que os observados por e Cortez (1996) e Bispo (2002) na área de Pegões (0,47, 0,16 e 0,16 g m-2 , respectivamente). Entre essas diferenças ressalta a inerente ao K, a qual poderá atribuirse a fontes (emissões) locais, dado que a área de estudo está inserida numa paisagem de rochas xistosas, cujos solos apresentam minerais (micas) com elevado teores de K. Os fluxos anuais das formas minerais de N (0,26 g m-2) são menores do que as estimadas noutras regiões do País (0,32-0,45 g m-2; Cortez, 1996), o que indica claramente que o local de estudo está ao abrigo de fontes poluidoras; porém, o fluxo anual de NOD (0,92 g m-2) foi cerca de 3,5 vezes superior ao das formas minerais.
Quadro 3 – Fluxos anuais de carbono (COD) e azoto (NOD) orgânicos dissolvidos e de nutrientes na PB, GO, ET e no gotejo líquido (GL)* e valores das correspondentes precipitações em 2001.
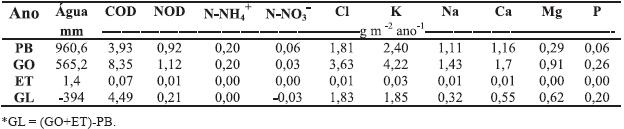
A devolução anual de Ca e Mg ao solo através gotejo (respectivamente de 0,55 e 0,62 g m-2) foi da ordem de grandeza da relatada para outras espécies florestais do País (Cortez, 1996; Bispo, 2002), ao passo que a de K (1,85 g m-2) foi muito mais elevada. A diferença respeitante ao K poder-se-á parcialmente atribuir, à maior precipitação na área do presente estudo, e que segundo esses mesmos autores é um factor com forte influência na quantidade de nutrientes lixiviados e/ou lavados da copa. Sublinha-se que a quantidade de K transferida anualmente para o solo via gotejo (cerca de 1,85 g m-2; Quadro 3) é superior à quantidade devolvida pela folhada (1,62 g m-2; Pereira, 2004), o que está em linha com o facto do ciclo do K incluir largas perdas da copa das árvores por gotejo (Fisher & Binkley, 2000) e este elemento não estar estruturalmente ligado em moléculas orgânicas. Os resultados do presente estudo indicam que o freixo (espécie caducifólia) - que não apresenta folhagem durante o período em que ocorre a maior parte da precipitação -, também apresenta forte influência na transferência de nutrientes para o solo via gotejo. No entanto, estudos mais detalhados, tendo em conta as componentes da sua biomassa, são necessários para avaliar a especificidade da espécie na translocação de nutrientes para o solo. Similarmente ao relatado para muitas espécies florestais em ecossistemas Mediterrâneos (Bellot & Escarré 1991; Cortez, 1996; Bispo, 2002), o fluxo anual de Ca pelo gotejo (0,55 g m-2), ao contrário do K, foi quase 10 vezes inferior ao fluxo devido à queda de folhada (5,1 g m-2; Pereira, 2004).
Ao contrário do observado para os catiões básicos, o fluxo de N-NO3 e N-NH4+ (Quadro 3) decorrente do gotejo (após passagem da precipitação bruta pela copa de freixo) foi negativo e nulo, respectivamente. O fluxo negativo de N-NO3-observado no presente estudo, foi também assinalado por Moreno et al. (2001) em Q. pyrenaica, por Rodrigo et al. (2003) em Q. ilex, por Avila et al. (2002) em Q. rotundifolia e por Moro et al. (1996) e Simões et al. (2009) em espécies arbustivas, em ecossistemas mediterrâneos com baixa disponibilidade de N. Para Rodrigo et al. (2003), o balanço negativo (absorção líquida na superfície das folhas e ramos) poderá resultar da absorção destes iões (N-NO3-e N-NH4+) pelos tecidos vegetais e, ainda, pela retenção destes pela flora epífita. Assim, os resultados do presente estudo sugerem que a folhagem do freixo poderá actuar como reservatório líquido para o N, indicando que o N proveniente da atmosfera é retido no ecossistema e contribui para a acumulação de N na vegetação e no solo, como sugerem Rodá et al. (2002) para florestas Mediterrâneas. A absorção de N pela folhagem de freixo também pode ser devida à baixa razão N/P (inferior a 10), a qual segundo Aerts & Chapin (2000), está longe da óptima da generalidade das plantas (15) e sugere limitação da nutrição em N.
Apesar das concentrações de quase todos os elementos analisados atingirem os valores mais elevados no escorrimento do tronco, à semelhança do observado por vários autores para outras espécies florestais (Bellot & Escarre, 1991; Moreno et al. 2001; Nunes et al., 2003; Rodrigo et al., 2003), as quantidades que atingem anualmente o solo são diminutas se forem expressas por unidade de área da projecção vertical da copa da árvore (Quadro 3). Porém, se considerarmos que esse escorrimento apenas influencia as áreas circunvizinhas ao tronco da árvore, as quantidades de nutrientes por unidade de área serão então muito maiores, variando segundo um modelo exponencial negativo com a distância ao tronco, como se pode observar na Figura 2. Assim, as quantidades de nutrientes transferidas para o solo na proximidade do tronco poderão ser muito superiores àquelas transferidas pelo gotejo; por exemplo, a quantidade de K devolvido anualmente ao solo, se admitirmos que se distribui por uma coroa circular de 25 cm de raio, pode atingir 4,7 g m-2, o que é muito superior à quantidade devolvida por gotejo (1,85 g m-2), considerando que a distribuição deste sob a copa é uniforme; porém, se considerarmos que a solução de escorrimento se distribui por uma coroa circular com 1,5 m de raio, as quantidades que atingem o solo são diminutas (Figura 2). A grande quantidade de nutrientes devolvidos ao solo na vizinhança do tronco é corroborada pela elevada concentração de K extraível observada no solo da proximidade do tronco do freixo (Pereira, 2004) e doutras espécies de sistemas tipo savana (Nunes, 2004; Belsky et al., 1989); com efeito, o primeiro autor observou teores de K e P extraíveis muito maiores a 25 cm do tronco (275 e 35 mg kg-1, respectivamente) do que no limite da projecção da copa da árvore (90 e 11 mg kg-1, respectivamente). Ou seja, o escorrimento do tronco contribui para o gradiente de fertilidade do solo da proximidade do tronco para a periferia da copa da árvore. Por fim, sublinha-se a elevada quantidade de COD que é transferida anualmente para o solo em redor do tronco da árvore (4,49 g m-2; Figura 2). Este fluxo de compostos orgânicos solúveis deverá estimular a actividade microbiana e influenciar o quociente metabólico e, por conseguinte, a dinâmica do carbono no solo, como foi observado por Pérez-Bejarano et al. (2008) para condições mediterrâneas.
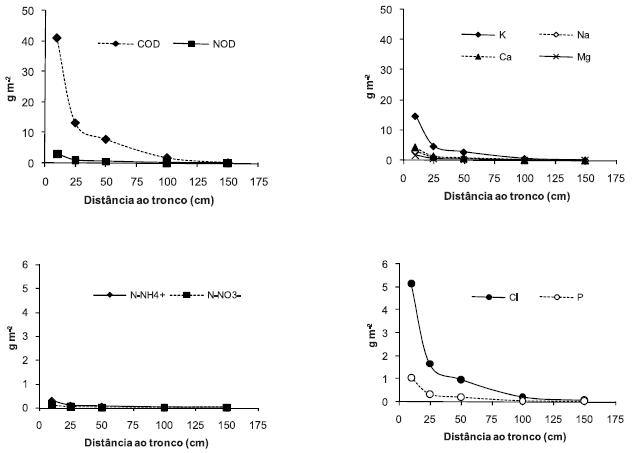
Figura 2 – Fluxos anuais médios de elementos pelo escorrimento do tronco, segundo um gradiente de distância em relação ao tronco (considerando uma coroa circular à volta do tronco da árvore com 0,1, 0,25, 0,50, 1,0 e 1,5 m de raio).
CONCLUSÕES
A deposição atmosférica (seca e húmida) foi mínima relativamente aos aniões considerados acidificantes, enquanto as correspondentes ao Ca e K são muito elevadas relativamente às determinadas noutros pontos do País. A passagem da precipitação através da copa do freixo conduziu, em geral, a um aumento dos valores de pH, dos teores de COD, NOD e de nutrientes, à excepção do NO3-, da solução de gotejo e escorrimento ao longo do tronco, em relação à PB, devido à lavagem e/ ou lixiviação dos tecidos vegetais. As elevadas quantidades de nutrientes restituídas ao solo através destas soluções poderão contribuir para a diferenciação das características químicas e físicas do solo sob copa. O presente estudo, também sugere que os inputs de catiões básicos estão relacionados com as condições do meio e que a quantidade de nutrientes devolvidos ao solo pela espécie caducifólia, podem ser superiores aos relatados para espécies perenifólias. Os resultados sugerem que a deposição atmosférica e o gotejo (e escorrimento do tronco) podem contribuir para minimizar processos de acidificação.
AGRADECIMENTOS
O presente estudo foi realizado no âmbito dos Projectos Influência das interacções solo -vegetação herbácea -árvore na valorização de sistemas agro florestais no Nordeste Transmontano (PAMAF 8047) e O freixo (Fraxinus angustifolia Vahl) como elemento valorizador dos sistemas agro-florestal e florestal no Nordeste Transmontano (AGRO 207). Os autores agradecem ao Prof. João Coutinho a realização das análises no laboratório de solos da UTAD, bem como às Engenheiras Ana Mafalda, Sónia Beito e Ana Pinto a colaboração nas tarefas de campo.
REFERÊNCIAS BIBLIOGRÁFICAS
Aber, J.D. & Melillo, J.M. (2001) -Terrestrial Ecosystems. Academia Press, London.
Aerts, R. & Chapin, F.S. (2000) -The mineral nutrition of wild plants revisited: A reevaluation of processes and patterns. Advances in Ecological Research 30: 1-67.
Attwill, P. & Leeper, G.W. (1987) -Forest Soils and Nutrient Cycles. Melbourne University Press, Carlton.
Ávila, A.; Rodrigo, A. & Roda, F. (2002) -Nitrogen circulation in a Mediterranean holm oak forest, La Castanya, Montseny, northeastern Spain. Hydrology and Earth Systems Sciences 6: 551-557
Bellot, J., & Escarré, A. (1991) Chemical characteristics and temporal variations of nutrients in throughfall and stemflow of three species in Mediterranean holm oak forest. Forest Ecology and Management 41: 125-135
Belsky, A.J.; Amundson, R.G.; Duxbury, J.M.; Riha, S.J.; Ali, A.R. & Mwonga, S.M. (1989) – The effects of trees on their physical, chemical and biologicalç environment in a semi-arid savanna in Kenya. Journal of Applied Ecology 26: 1005-1024.
Bispo, R.M.B. (2002) -Meteorologia da Camada Limite e Deposição Atmosférica numa Superfície Vegetal (Eucalyptus globulus). Dissertação de Doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Lisboa.
Casartelli, M.R.; Mirlean, N.; Peralba, M.C.; Barrionuevo, S.; Gómez-Rey, M.X. & Madeira, M. (2008) -An assessment of the chemical composition of precipitation and throughfall in rural-industrial gradients in wet subtropics (Southern Brazil). Environmental Monitoring and Assessment 144, 1: 105-116.
Cortez, N.R.S. (1996) -Compartimentos e Ciclos de Nutrientes em Plantações de Eucalyptus globulus Labill. ssp. globulus e Pinus pinaster Aiton. Dissertação de Doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Lisboa.
Cortez, N.; Nunes, J. & Madeira, M. (2004) -Efeito do escorrimento ao longo dos troncos de Eucalyptus globulus, Pinus pinaster e Quercus rotundifolia no padrão de variação espacial de propriedades do solo. Revista de Ciências Agrárias 27, 1: 470-480. [ Links ]
FAO (1988) - FAO - UNESCO Soil Map of the World, Revised Legend, with corrections. World Resources Report 60, FAO, Rome (Reprinted as Technical Paper 20, ISRIC, Wageningen, 1994).
Fisher, R. F. & Binkley, D. (2000) – Ecology and Management of Forest Soils (Third Edition). John Wiley & Sons, Inc. New York.
Gauquelin, T.; Fromard, F.; Badri, W. & Dagnac, J. (1992) - Apports déléments minéraux au sol par lintermédiaire de la litère, des pluies et des pluviolessivats dans un peuplement à genévrier (Juniperus thurifera L) du Haut Atlas occidental (Maroc). Annals of Forest Science 49: 599-614.
Gersper, P.L. & Holowaychuk, N. (1970) - Effects of stemflow water on a Miami Soil near a beech tree: II. Chemical properties. Soil Science Society American Proceedings 34:786-794.
Houba, V.J.G.; Novozamski, I. & Tenminghoff, E. (1994) -Soil Analysis Procedures. Department of Soil Science and Plant Nutrition, Wageningen Agricultural University, The Netherlands.
INMG (1991) - Normais Climatológicas da Região de «Trás-os-Montes e Alto Douro e Beira Interior», Correspondentes a 1951-1980. O Clima de Portugal. Fascículo XLIX, vol. 3 -3ª Região. Instituto Nacional de Meteorologia e Geofísica, Lisboa.
Levia, D.F. & Herwitz, S.R. (2002) -Winter chemical leaching from deciduous tree branches as a function of branch inclination angle in central Massachusetts. Hydrological Processes 16: 2867-2879.
Ludwig, F. (2001) – Tree-Grass Interactions on an East African Savanna: The Effects of Competition, Facilitation and Hydraulic Lift. Tropical Resource Management Papers, No. 39.
Marschner, H. (1995) - Mineral Nutrition of Higher Plants. Academic Press, London.
Meiwes, K.J.; Merino, A. & Beese, F.O. (1998) -Chemical composition of throughfall, soil water, leaves and leaf litter in a beech forest receiving long term application of ammonium sulphate. Plant and Soil 201: 217-230.
Moreno, G.; Gallardo, J.F. & Bussotti, F. (2001) -Canopy modification of atmospheric deposition in oligotrophic Quercus pyrenaica forests of an unpolluted region (central-western Spain). Forest Ecology and Management 149: 47-60.
Moro, M.J.; Domingo, F. & Escarré, A. (1996) -Organic matter and nitrogen cycles in a pine afforested catchment with a shrub layer of Adenocarpus decorticans and Cistus laurifolius in south-eastern Spain. Annals of Botany 78: 675-685
Nunes, J.M.S.D. (2004) – Interacção Solo – Árvore Isolada em Montados de Azinho (Quercus rotundifolia Lam.) Dissertação de Doutoramento em Biologia. Universidade de Évora, Évora.
Nunes, J.; Madeira, M. & Gazarini, L. (2003) - Fluxos de nutrientes em árvores de montado de Quercus rotundifolia Lam., no Sul de Portugal. Revista de Ciências Agrárias 28:328-342 [ Links ]
Parker, G.G. (1983) -throughfall and stemflow in the forest nutrient cycle. Advances in Ecological Research 13: 57-133.
Pereira, E.L. (2004) - Influência do Freixo no Microclima, nas Características do Solo e Disponibilidade de Nutrientes, e na Vegetação Herbácea de Lameiros do Nordeste de Portugal. Dissertação de Doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Lisboa.
Pereira, E.; Abreu, F.G. & Madeira, M. (2007) – Influência de uma árvore isolada de freixo (Fraxinus angustifolia Vahl) no microclima num lameiro em Trás-os-Montes. Revista de Ciências Agrárias 30: 233-245. [ Links ]
Pereira, E.; Ribeiro, A. & Castro, P. (2000) -Carta Geológica de Portugal na escala 1:200 000, Folha 2. Instituto Geológico e Mineiro. Ministério da Economia, Lisboa.
Pérez-Bejarano, A.; Mataix-Solera, J.; Zornoza, R.; Guerrero, C.; Arcenegui, V.; Mataix-Beneyto, J. & Cano-Amat, S. (2008) -Influence of plant species on physical, chemical and biological soil properties in a Mediterranean forest soil. European Journal of Forest Research. DOI 10.1007/s10342-008-0246-2. Disponível em: <http://www.springerlink.com/content/w216k84834448457/> (acesso em: 13 de Maio de 2009).
Portela, E. & Pires, A.L. (1995) -Nutrient deposition and leaching by rainwater in low and intensively managed chestnut groves. In: Proceedings of the Conference on Erosion and Land Degradation in the Mediterranean, Universidade de Aveiro, 14 a 18 de Junho, pp. 307-317.
Rodá, F.; Avila, A. & Rodrigo, A. (2002) -Nitrogen deposition in Mediterranean forests. Environmental Pollution 118: 205-213.
Rodrigo, A.; Ávila, A. & Rodá, F. (2003) -The chemistry of precipitation, throughfall and stemflow in two holm oak (Quercus ilex L.) forests under a contrasted pollution environment in NE Spain. The Science of the Total Environment 305: 195-205.
Seiler, J. & Matzner, E. (1995) -Spatial variability of throughfall chemistry and selected soil properties as influenced by stem distance in a mature Norway spruce (Picea abies, Karst.). Plant and Soil 176: 139-147.
Simões, M. P.; Madeira, M. & Gazarini, L. (2009) - Ability of Cistus L. shrubs to promote soil rehabilitation in extensive oak woodlands of Mediterranean areas. Plant and Soil DOI 10.1007/s11104-009-9934-z Disponível em: <http://www.springerlink.com/content/r94554k10v401165/fulltext.pdf.> (acesso em: 12 de Maio 2009).
1 Superior Agrária de Bragança, Apartado 172, 5301-855 Bragança, Portugal – Tel: (+351) 273 303 384 – Fax: (+351) 273 325 405; e-mail: epereira@ipb.pt;
2 Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Tapada de Ajuda, 1349-017, Lisboa – Tel: (+351) 213 653 100; e-mail: mavmadeira@isa.utl.pt
Recepção/Reception: 2009.06.08
Aceitação/Acception: 2009.07.08

