










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAnálise Psicológica
versão impressa ISSN 0870-8231
Aná. Psicológica v.26 n.3 Lisboa jul. 2008
Dominância e atracção social num grupode macacos-verdes (Cercopithecus aethiops) em cativeiro
João R. Daniel (*)
António J. Santos (**)
Luís Vicente (***)
RESUMO
Dados de interacções de catagem, recolhidos durante quatro meses de observações directas, de um grupo de macacos-vervet (Cercopithecus aethiops) em cativeiro, no Jardim Zoológico de Lisboa, foram utilizados para testar o modelo clássico de Seyfarth (1977) de atracção por indivíduos dominantes. Cada indivíduo foi observado um total de 10.5 horas, através de amostragens focais. Todas as ocorrências de comportamentos agonísticos foram também registadas. A análise dos resultados confirmou que nas fêmeas a catagem é dirigida para cima na hierarquia, apesar de a troca de catagem por apoio agonístico não ser o princípio subjacente a esta atracção. Considerando a teoria dos mercados biológicos, o facto de as díades serem altamente não recíprocas em termos do investimento feito na catagem, parece sugerir que outras comodidades, para além do apoio agonístico, estão a ser trocadas.
Palavras-chave: Macacos-verdes, catagem, atracção social, apoio agonístico.
ABSTRACT
Data of grooming interactions, collected through four months of direct observation of a captive group of vervet monkeys (Cercopithecus aethiops), in Lisbon Zoo, were used to test Seyfarth’s (1977) classical model of rank related attractiveness. Each individual was observed for a total of 10.5 hours by means of focal sampling. All occurrences of agonistic behaviours were also recorded. Analyses confirmed that in females grooming is directed up the hierarchy, despite the interchange of grooming for support not being the underlying principle of this attraction. Considering biological markets theory, the fact that groom dyads tend to be highly non-reciprocal may actually suggests that other commodities, besides support, are being traded.
Key words: Vervet monkeys, grooming, social attraction, agonistic support.
INTRODUÇÃO
A catagem é de longe o comportamento mais frequentemente estudado em primatas não humanos, uma vez que para além da sua função higiénica, há muito que é enfatizado o seu papel na formação de laços sociais, especialmente em fêmeas, dado que entre machos a sua ocorrência é muito menos frequente. Desempenhando a catagem um papel fundamental nas estratégias sociais de primatas, muitos dos modelos sobre cooperação nestas espécies procuram explicar a sua distribuição não aleatória, pelos diferentes membros do grupo.
Um dos modelos mais influentes foi proposto por Seyfarth (1977), depois de verificar que em várias espécies de macacos e babuínos, as fêmeas de elevado estatuto de dominância recebiam mais catagem que as de baixo estatuto, ao mesmo tempo que a maior parte da catagem ocorria entre fêmeas de posições hierárquicas adjacentes. O modelo de Seyfarth (1977) baseia-se na assumpção principal de que as fêmeas trocam catagem por apoio em confrontos agonísticos futuros. Dado que as fêmeas dominantes são as parceiras mais valiosas durante confrontos agonísticos (i.e. possuem maior probabilidade de vencer uma disputa), as outras fêmeas deverão ser atraídas por elas, com o intuito de maximizar os benefícios das relações sociais. Dado que cada fêmea competirá por parceiras o mais dominantes possíveis, o acesso a estas não estará ao alcance de todas, fazendo com que cada fêmea despenda mais tempo a catar fêmeas de posições hierárquicas adjacentes (Seyfarth, 1977).
A distribuição preferencial de catagem por fêmeas dominantes tem sido testada especialmente em primatas do Velho Mundo. Correlações significativas entre estatuto de dominância e catagem recebida foram encontradas em algumas espécies de primatas (Fairbanks, 1980; Hemelrijk & Ek, 1991; Matheson & Bernstein, 2000; Nakamichi, 2003; Payne et al., 2003; Rowell, 1971; Schino et al., 2003; Seyfarth, 1980; de Waal & Luttrell, 1986; Watts, 2000), não sendo, no entanto, uma tendência geral em todas elas (Bentley-Condit & Smith, 1999; Cooper & Bernstein, 2000; Franz, 1999; Leinfelder et al., 2001; Silk et al., 1999).
Infelizmente, alguns destes estudos em que foram encontradas associações significativas, não levaram em conta o papel das relações de parentesco na distribuição da catagem. Muitas destas espécies formam linhas matriarcais, em que as descendentes adquirem posições hierárquicas imediatamente inferiores às das suas progenitoras, pelo que, a aparente atracção por indivíduos dominantes pode não ser mais que o investimento em relações de parentesco.
Como inicialmente sugerido por Seyfarth (1977), um dos benefícios principais destas relações seria a formação de alianças agonísticas. No entanto, correlações significativas entre a catagem e a formação destas alianças, apesar de se terem observado em alguns grupos (Hemelrijk, 1994; Hemelrijk & Ek, 1991; Seyfarth, 1980; Seyfarth & Cheney, 1984), não foram encontradas noutros (Cooper & Bernstein, 2000; Fairbanks, 1980; Gust, 1995; Leinfelder et al., 2001; Matheson & Bernstein, 2000; Silk et al., 2004; de Waal & Luttrell, 1986), sugerindo que o funcionamento da catagem como um meio de fortalecer alianças agonísticas entre fêmeas, não pode ser tomado com uma explicação geral para todos os primatas (Barrett & Henzi, 2002).
Expandindo as ideias iniciais de Seyfarth (1977), Henzi e Barrett (1999) abordaram o estudo da função social da catagem através de um ângulo mais vasto, ao considerar a catagem como um ‘bem’, que pode ser trocado por ele próprio, ou por um conjunto de outros ‘bens’, que não exclusivamente o apoio agonístico (e.g. comida: de Waal, 1997; tolerância em locais de alimentação: Barrett et al., 2002; Pastor-Nieto, 2001; acesso a crias: Bentley-Condit & Smith, 1999; di Biteti, 1997; Henzi & Barrett, 2002; Muroyama, 1994; Silk, 1999; cuidados alo-parentais: Lazaro-Perea et al., 2004). Estes autores, baseando-se na teoria dos mercados biológicos (Noë & Hammerstein, 1994, 1995), sugerem que as interacções de catagem entre fêmeas reflectem um mercado dinâmico de trocas entre parceiros comerciais, no qual, o preço dos bens é influenciado pelas leis da oferta e da procura, e pelo estatuto de dominância dos indivíduos, uma vez que este regula o acesso preferencial aos recursos (Barrett et al., 1999; Barrett & Henzi, 2002).
Apesar das diferenças, o modelo de Seyfarth (1977) pode ser visto como uma forma simplificada de um modelo de mercado (troca de catagem por alianças), uma vez que este assenta igualmente na competição por parceiros de diferente valor (Barrett & Henzi, 2005). Contudo, o modelo de Seyfarth é inteiramente estático, porque assume que o valor das fêmeas de elevado estatuto mantém-se constante ao longo do tempo, independentemente das variações no contexto socio-ecológico (Barrett & Henzi, 2005).
Esta perspectiva de mercado biológico assume que quando a catagem não é trocada por outros bens, os indivíduos formam relações recíprocas, pelos benefícios inerentes à própria catagem (e.g. remoção de parasitas, redução de tensão), recorrendo à reciprocação imediata (Barrett et al., 1999). Assim, segundo este modelo, os padrões temporais da catagem poderão ser usados para distinguir duas classes de parceiros: recíprocos ou permutadores (Henzi & Barrett, 1999; Leinfelder et al., 2001). Apesar de tudo, esta distinção pode não ser tão linear quanto isso, uma vez que as relações inter-individuais são melhor caracterizadas por uma mistura complexa de permuta e reciprocidade (Schino et al., 2003), e pelo facto de nem todas as interacções serem obrigatoriamente influenciadas por uma lógica de mercado (e.g. investimento parental) (Barrett & Henzi, 2005).
Contudo, para se comprovar a existência de um mercado biológico, é necessário que as condições ecológicas variem suficientemente, de modo a que haja um impacto significativo no regime competitivo, e consequente alteração no investimento nas relações sociais (Barrett & Henzi, 2005). No entanto, mesmo na ausência de oscilações nas leis de oferta e de procura dos mercados biológicos, podem ser feitas um conjunto de predições assumindo que a catagem é de facto um bem de mercado, que pode ser trocado por outros bens, e que o preço a pagar por esses bens é influenciado pela capacidade diferencial dos indivíduos de controlarem recursos. Neste sentido, Leinfelder et al. (2001) tentaram incorporar no modelo de Seyfarth (1977), um conjunto de assumpções mais gerais que lhe permitem ser aplicado a diferentes tipos de hierarquias de dominância.
Então, segundo Leinfelder et al. (2001) e assumindo um modelo de troca de catagem num grupo de fêmeas em que existe um elevado declive (assimetria) nas relações de dominância, podem ser feitas as seguintes predições: 1) atracção por fêmeas dominantes (fêmeas dominantes recebem mais catagem, fêmeas subordinadas realizam mais catagem, fêmea subordinada da díade responsável pela maior parte da catagem); 2) mais catagem entre díades adjacentes; 3) maior reciprocidade quanto menor a distância em termos de estatuto de dominância; e 4) quanto maior a distância em termos de estatuto, maior a contribuição do membro subordinado de cada díade. Se por outro lado, assumirmos que as relações de dominância são mais igualitárias e existem poucos bens pelos quais se possa trocar a catagem, então podemos esperar: 5) elevada reciprocidade de catagem; e 6) maior reciprocidade em díades onde a catagem ocorre mais frequentemente.
Contudo, Leinfelder et al. (2001) ao realizarem as suas predições ignoraram parte das principais diferenças entre o modelo de Seyfarth (1977) e o modelo de mercado biológico (Barrett & Henzi, 2005). O primeiro modelo assume que as fêmeas ao acederem a parceiros dominantes, excluem automaticamente as fêmeas de estatuto inferior de acederem às mesmas fêmeas que elas (i.e. as fêmeas dominantes são as primeiras a escolher e automaticamente impedem as restantes de oferecerem uma melhor proposta). O segundo modelo, por seu lado, considera que fêmeas que oferecem um produto de melhor qualidade (ou pedem um preço mais baixo) têm mais sucesso no mercado. Em termos práticos, esta diferença faz com que as predições de Leinfelder et al. (2001), nomeadamente as que dizem respeito a maior catagem e maior reciprocidade entre díades adjacentes, não se apliquem necessariamente em todos os casos.
Apesar de, em termos teóricos, o declive (assimetria) das relações de dominância ser há muito um conceito familiar no campo da primatologia (e.g. despotismo como sinónimo de relações de dominância lineares com grandes assimetrias em termos de poder), só recentemente foi operacionalizada por de Vries et al. (2006). A medida proposta permite quantificar as diferenças absolutas entre indivíduos, quanto ao seu sucesso em encontros agonísticos. Partindo desta medida, e tendo por base as interacções de catagem num grupo cativo de macacos-verdes (Cercopithecus aethiops) testámos cada uma das predições de Leinfelder et al. (2001) acima enunciadas, verificando até que ponto os resultados obtidos se enquadram mais no tipo de competição prevista pelo modelo de atracção de Seyfarth (1977), ou pelo modelo dos mercados biológicos (Barrett & Henzi, 2005). Tendo em conta que, em macacos-verdes as fêmeas formam o núcleo central do grupo e as filhas assumem uma posição hierárquica semelhante à das mães (Melnick & Pearl, 1987), prevemos a existência de um mercado de catagem entre estas, podendo este estar, ou não, directamente relacionado com a formação de alianças agonísticas, tal como previsto por Seyfarth (1977).
Resultados prévios em macacos-verdes obtidos por Lynn Fairbanks apoiam a hipótese de atracção e competição por fêmeas de elevado estatuto (Fairbanks, 1980), apesar de não sido encontrada uma relação causal entre catagem e formação de alianças agonísticas. Seyfarth (1980), por outro lado, encontrou evidências de que de facto, elevadas frequências de catagem se encontravam signitivamente correlacionadas com a formação de alianças. No entanto, uma re-análise dos dados de Seyfarth (1980) por Hemelrijk (1990) sugere que, a troca entre catagem e apoio agonístico é um subproduto da correlação de ambas as variáveis com o estatuto de dominância (i.e. indivíduos dominantes tendem a receber mais catagem e a envolverem-se mais frequentemente na formação de alianças). O facto de no presente estudo, as fêmeas adultas do grupo não serem aparentadas entre si, permite avaliar a existência fenómenos de atracção independentemente do eventual efeito das relações de parentesco.
MÉTODO
Sujeitos
Os macacos-verdes encontravam-se alojados numa jaula exterior de aproximadamente 300m3, no Jardim Zoológico de Lisboa. O espaço possuía uma forma octogonal, tecto parcialmente aberto e chão de cimento, contendo diversas estruturas de madeira que forneciam sombra e superfícies adicionais para trepar e sentar. Tinha ainda três pequenos abrigos onde os animais podiam dormir e proteger-se das condições climáticas. A jaula era limpa diariamente, e os macacos alimentados duas vezes por dia.
O grupo era constituído por três machos (um adulto >10 anos – AM1, dois subadutlos >4 anos – SAM1-2) e sete fêmeas (cinco adultas >5 anos – AF1-5 e duas juvenis – 3 e 2 anos JF1-2). Nenhum dos adultos nasceu no zoo, entrando no grupo em alturas distintas. Os restantes animais já nasceram no zoo de Lisboa (SAM1-2, JF1-2), mas somente a fêmea mais nova (JF2) possuía a mãe presente (contudo os registos do zoo não são conclusivos quanto a qual seria o par mãe-cria)
Recolha de dados
Todos os dados foram recolhidos pelo primeiro autor de Março a Julho de 2003, entre as 10:00H e as 18:00H. Cada animal foi sujeito a 126 amostragens focais (Altmann, 1974) de cinco minutos. Durante estas amostragens, todas as ocorrências dos seguintes comportamentos foram registadas: 1) aproximação/evitamento: um animal afasta-se rapidamente de outro que potencialmente se aproxima, 2) catagem: um animal trata a pele/pelagem de outro (a duração destas interacções foi registada ao segundo), 3) agressão: ameaçar, bater, puxar, morder ou perseguir outro animal, 4) apoio agonístico: um animal intervém num confronto a favor de um dos intervenientes. As descrições comportamentais são baseadas em Fairbanks (1980), Hector et al. (1989) e Raleigh e McGuire (1989). Para cada ocorrência destes comportamentos, a direcção e a identidade dos sujeitos que interagiram com o sujeito focal foram registados. Uma vez que episódios agonísticos ocorrem pouco frequentemente, estes (bem como eventuais apoios agonísticos) foram igualmente registados segundo a amostragem de todas as ocorrências (Altmann, 1974). A ordem de amostragem dos indivíduos foi aleatória, sendo que o mesmo indivíduo não foi observado uma segunda vez, sem que todos os outros o tivessem sido primeiro.
Análise dos dados
Hierarquia de dominância
Através do resultado das interacções de aproximação/evitamento foi construída uma hierarquia de dominância baseada no método proposto por David (1987, 1988), com a correcção de de Vries et al. (2006). Este método atribui um valor de dominância para cada indivíduo (David score, DS), no intervalo de -N(N-1)/2 (valor mínimo possível, em que N representa o número de sujeitos) a N(N-1)/2 (valor máximo possível). A grande vantagem deste procedimento é que valoriza as vitórias diferentemente, atribuindo maior peso a vitórias sobre indivíduos dominantes, do que a vitórias sobre indivíduos que perdem a maioria dos seus confrontos. Igualmente, derrotas provocadas por indivíduos subordinados, têm maior peso do que derrotas em confrontos com indivíduos dominantes.
Para se obter uma medida de declive das relações de dominância, que varie entre 0 e 1, é necessário normalizar os valores de DS (NormDS) utilizando a seguinte expressão: NormDS = {DS + MaxDS(N)} / N = {DS + N(N-1) / 2} / N (de Vries et al., 2006).
Os valores de NormDS encontrar-se-ão definidos no intervalo entre 0 e N-1. Colocando os indivíduos por ordem de dominância no eixo dos X (1 para o indivíduo mais dominante, N para o menos dominante) e os respectivos valores NormDS no eixo dos Y, o declive das relações de dominância será dado pelo valor absoluto do declive da recta de mínimos quadrados, ajustada ao conjunto de pontos (de Vries et al., 2006). Valores próximos de 1 representam relações de dominância altamente assimétricas, enquanto valores próximos de 0 correspondem a relações de dominância mais igualitárias.
Foi ainda calculado um índice de linearidade para a hierarquia de dominância (de Vries, 1995). Este índice mede o grau de transitividade das relações de dominância. Uma hierarquia é completamente linear se, e só se, verificarem as seguintes condições: 1) para cada díade A e B existente no grupo, ou A domina B ou B domina A; e 2) para cada trio de indivíduos no grupo A, B e C, se A domina B, B domina C, então A domina C.
O cálculo dos David scores, declive e linearidade da hierarquia de dominância foi efectuado tendo por base a matriz diádica contendo todos os elementos do grupo (n=10, 3 machos, 7 fêmeas).
Catagem
Para determinar se a distribuição da catagem se encontrava relacionada com o estatuto de dominância, os tempos totais de catagem, recebida e efectuada, por cada uma das fêmeas foi correlacionada (coeficiente de correlação de Spearman) com o respectivo DS. Adicionalmente, recorrendo ao teste binomial, comparámos a proporção de díades em que a catagem foi maioritariamente (>50%) realizada pela fêmea subordinada, com a proporção de díades em que esta foi maioritariamente efectuada pela fêmea dominante da díade.
Para determinar se os tempos de catagem foram maiores em díades adjacentes, comparámos o tempo médio de catagem, que cada fêmea dedicou às fêmeas imediatamente acima ou abaixo na hierarquia (somente abaixo no caso da fêmea mais dominante e somente abaixo no caso da fêmea mais subordinada), com o tempo médio dedicado às restantes, utilizando o teste de Wilcoxon para amostras emparelhadas. Um procedimento semelhante foi utilizado para avaliar se a reciprocidade foi maior nestas díades. A reciprocidade da catagem entre as díades foi calculada segundo a seguinte expressão:
R = 1 - ¦ (Cab-Cba)/(Cab+Cba) ¦
Um valor de R=1 representa reciprocidade total, e um valor de 0 representa que um dos membros da díade foi responsável por toda a catagem.
Para cada uma das díades foi ainda correlacionada (coeficiente de correlação de Spearman) o valor da diferença dos DS (DS dominante – DS subordinada) com a reciprocidade observada e com a contribuição relativa da fêmea de estatuto mais baixo.
Finalmente, nas díades em que se observaram ocorrências de apoio agonístico, os tempos de catagem foram comparados com um valor médio representativo de todas as díades, onde não se verificaram ocorrências deste comportamento (teste de Wilcoxon para amostras emparelhadas).
RESULTADOS
Em média, cada indivíduo passou 9.43 ± 9.00 min/h (média ± desvio padrão) a catar outros membros do grupo, com as fêmeas adultas a dedicarem mais tempo à catagem que os restantes indivíduos (fêmeas adultas = 16.12 ± 7.88 min/h; fêmeas juvenis = 5.59 ± 2.38 min/h; machos subadultos = 1.22 ± 0.84 min/h; macho adulto = 0.13 min/h). Por outro lado, as fêmeas juvenis e o macho adulto (dominante) receberam mais catagem que os restantes elementos do grupo (fêmeas juvenis = 14.04 ± 0.51 min/h; macho adulto = 11.75 min/h; fêmeas adultas = 8.68 ± 7.12 min/h; machos subadultos = 5.56 ± 4.22 min/h; Figura 1).
Catagem realizada e recebida pelas fêmeas de acordo com o seu estatuto de dominância
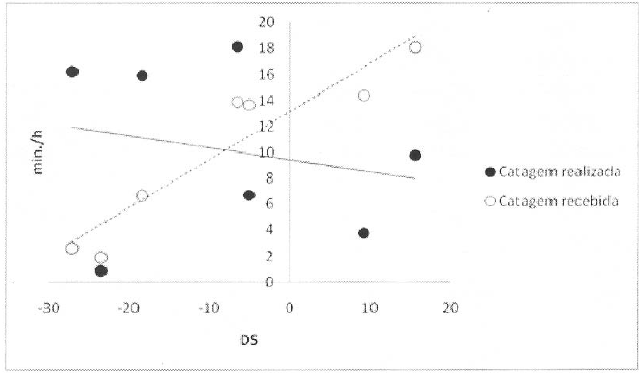
Legenda: DS – David score.
A hierarquia de dominância obtida apresentava um declive (s=0.608, p<0.001, 1000 simulações) e uma linearidade significativos (h’=0.899, p<0.001, 1000 simulações).
Não foi encontrada uma correlação significativa entre o tempo que cada fêmea despendeu a catar as restantes e o seu estatuto de dominância (coefeciente de correlação Spearman: n=7, r=-0.214, p=0.322, unilateral). Contudo, as fêmeas dominantes foram o alvo da maior parte da catagem das suas companheiras (coefeciente de correlação Spearman: n=7, r=0.929, p=0.001, unilateral; Figura 1). Em 18 das 21 díades existentes, mais catagem foi realizada pela fêmea de menor estatuto (teste binomial: p=0.01), com 70.9% da catagem a ser dirigida para cima na hierarquia.
De um modo geral, as fêmeas não forneceram mais catagem aquelas de posições adjacentes (adjacentes: 2.60 ± 2.37 min/h; não-adjacentes: 1.46 ± 1.38 min/h; Wilcoxon: n=7, p=0.188, unilateral; Figura 2). A ausência de significância estatística deve-se aos resultados das duas fêmeas mais subordinadas, uma vez todas as outras os tempos médios de catagem, dedicados a fêmeas de posições adjacentes são superiores (Wilcoxon: n=5, p=0.031, unilateral). No entanto, apesar de as médias serem superiores nestas cinco fêmeas, nem sempre, para as quatro fêmeas que possuem parceiras imediatamente acima e abaixo na hierarquia (fêmea dominante excluída), as duas fêmeas adjacentes são ambas as suas parceiras preferidas.
Tempos de catagem médios dedicados às fêmeas de posições hierárquicas adjacentes e não-adjacentes
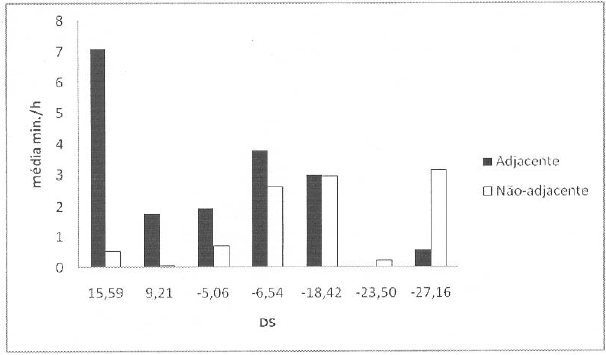
Legenda: DS – David score.
Na Figura 3 podemos observar que todas as fêmeas, à excepção das fêmeas AF3 e AF4 (DS -18.4 e -23.5 respectivamente), possuem a fêmea dominante como um dos parceiros preferidos de catagem. A fêmea subordinada do grupo é ainda a segunda que mais catagem realiza à fêmea dominante. A fêmea adulta (AF4) situada na penúltima posição da hierarquia apresentou um padrão de catagem muito diferente das suas companheiras, uma vez que para além de realizar muito menos catagem que as restantes fêmeas (ver Figura 1), dirigiu-a preferencialmente para dois dos machos do grupo (AM1 e SAM1; Figura 3).
Parceiros preferidos de catagem

Legenda: – macho; O– fêmea; os David scores de cada indivíduo encontram-se no interior dos quadrados/círculos; o sentido da seta A →B indica que o indivíduo A catou o B 1.5 vezes mais que o seu valor médio para todos os sujeitos.
A reciprocidade não foi também significativamente maior nestas díades (adjacentes: 0.37 ± 0.35; não-adjacentes: 0.30 ± 0.30; Wilcoxon: n=7, p=0.148, unilateral), e em média 67.8 ± 31.1% de toda a catagem entre díades não foi reciprocada. No entanto, foi encontrada uma tendência não significativa (p<0.1) para a correlação entre a distância entre as posições hierárquica e a quantidade de catagem reciprocada (coefeciente de correlação Spearman: n=21, r=-0.345, p=0.063, unilateral), com as díades mais distantes a reciprocarem menos catagem. Esta menor reciprocidade é acompanhada por maiores contribuições relativas da fêmea subordinada da díade (coefeciente de correlação Spearman: n=21, r=0.590, p=0.002, unilateral; Figura 4).
Reciprocidade de catagem e proporção da contribuição da fêmea subordinada para o total de catagem observado na díade, consoante a distância entre as posições hierárquicas das duas fêmeas
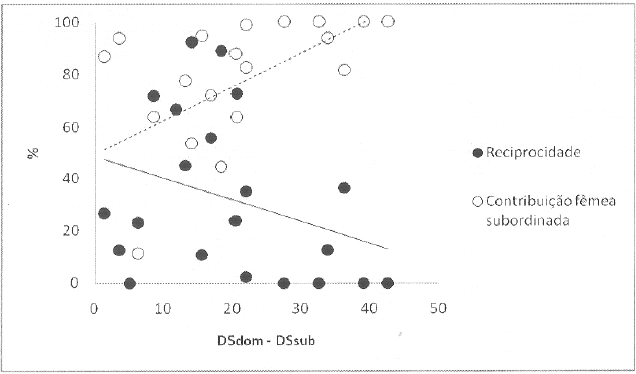
Legenda: DS – David score; dom – dominante; sub – subordinada.
Durante o período de estudo foram observados 105 conflitos (~0.9 conflictos/h), 73 dos quais foram diádicos, 20 triádicos e 12 poliádicos, com os apoiantes a intervirem sempre em favor do agressor. A fêmea juvenil mais velha (JF1) recebeu mais de metade de todo o apoio concedido (23/45). A outra fêmea juvenil aparece (JF2) em segundo lugar (7/45), e a fêmea dominante (AF1) em terceiro (5/45). A fêmea (AF1=12/45) e o macho dominante (AM1=8/45), juntamente com as fêmeas juvenis (JF1=13/45, JF2=8/45), são praticamente responsáveis pela formação de todas as alianças (41/45) (Figura 5).
Ocorrências de apoio agonístico
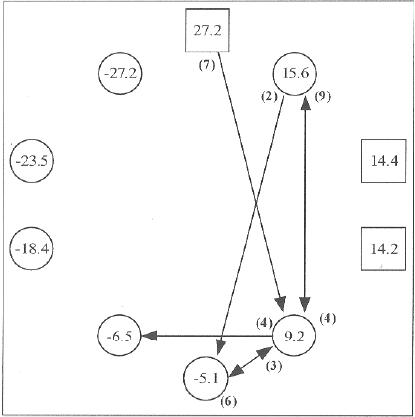
Legenda: – macho; O– fêmea; os David scores de cada indivíduo encontram-se no interior dos quadrados/círculos; o sentido da seta A →B representa apoio agonístico concedido por A a B, e o número de ocorrências encontra-se entre parêntesis junto ao indivíduo A; somente alianças verificadas mais de uma vez estão representadas.
Em 4 das 13 díades (AF1-JF1, AF1-JF2, AF2-JF1, JF1-JF2) em que se observou a formação de alianças (machos e fêmeas), os indivíduos reciprocaram estas alianças em alguma extensão. Em 8 das 9 díades restantes, um indivíduo intervém a favor do outro uma única vez.
Díades de fêmeas em que se registaram ocorrências de apoio agonístico (n=7), cataram-se mais que a díade média de fêmeas, onde não se observaram alianças (apoiantes: 4.99 ± 2.77 min/h; díade média:
2.59 ± 2.67 min/h; Wilcoxon: n=7, p=0.039, unilateral). Contudo, somente num dos 105 conflitos observados, é que uma fêmea adulta apoiou outra, e nunca duas fêmeas adultas foram vistas juntas a apoiar um terceiro indivíduo. Fica assim de lado, qualquer possibilidade de troca de catagem por apoio agonístico em fêmeas adultas.
DISCUSSÃO
Baseado nos elevados valores de linearidade e declive obtidos para a hierarquia de dominância de macacos-verdes (altamente transitiva e assimétrica), o modelo de troca de catagem prevê uma permuta potencial de catagem por outros benefícios relacionados com o estatuto hierárquico das fêmeas, cujo reflexo será uma distribuição da catagem claramente influenciada pela posição hierárquica destas. Os resultados deste estudo apoiam a grande maioria das predições de Leinfelder et al. (2001), contudo o ajustamento a estas não é perfeito.
Apesar de a grande maioria da catagem ser dirigida para cima na hierarquia, as fêmeas subordinadas não dedicaram, de um modo geral, mais tempo a esta actividade do que as fêmeas dominantes (Figura 1). Este último resultado deve-se especialmente ao facto de uma das fêmeas mais subordinadas (AF4) realizar muito menos catagem que qualquer uma das suas companheiras, e também à existência de um efeito da idade no tempo dedicado à catagem, com as fêmeas juvenis a investirem aproximadamente 1/3 do tempo das fêmeas adultas. A existência de maiores tempos de catagem em díades adjacentes não ficou também claramente demonstrada. Apesar de nas cinco primeiras fêmeas da hierarquia, os tempos médios de catagem dedicados a fêmeas de posições hierárquicas adjacentes serem superiores aos tempos dedicados às restantes fêmeas, esta diferença é conseguida especialmente à custa do investimento na fêmea imediatamente acima na hierarquia. Estes resultados ajudam igualmente a explicar a não existência de uma maior reciprocidade em díades adjacentes. Contudo, tal como previsto, quanto maior a distância em termos de estatuto hierárquico, menor a reciprocidade observada, e maior a contribuição da fêmea subordinada, para o total de catagem da díade.
Uma das explicações, para a não confirmação de todas as predições de Leinfelder et al. (2001), poderá residir no facto de grupos pequenos terem maior tendência a ser influenciados por ruído, gerado por diferenças individuais que não o estatuto de dominância (Sambrook et al., 1995). Outra explicação poderá ser a de que, a assumpção de que as fêmeas dominantes escolhem primeiro as suas parceiras impedindo o acesso a estas por partes das fêmeas subordinadas, não se verifique. Contudo, a hipótese alternativa, de que as fêmeas entram num mercado de licitações, oferecendo diferentes propostas, não ficou claramente demonstrada, uma vez que a adjacência de estatuto, parece ainda assim ter um papel relativamente importante. No entanto, existe alguma margem de manobra para considerar uma actuação conjunta dos dois tipos de mecanismo.
Apesar de podermos aceitar o modelo de troca de catagem, independentemente de como se processa a competição por parceiros, este não teve como consequência a formação de alianças agonísticas, nomeadamente no que diz respeito às fêmeas adultas, o que está de acordo como os resultados obtidos por Fairbanks (1980). As fêmeas adultas investiram bastante tempo nas relações com outras fêmeas adultas (e em menor grau com os machos), sem que isso se traduzisse em apoio concedido.
A maioria das coligações observadas foi realizada dentro de um núcleo restrito, constituído pela fêmea e macho dominante, juntamente com as duas fêmeas juvenis, com a maioria das alianças a serem realizadas a favor das fêmeas juvenis, ou a contarem com a sua participação. Pelo facto de as relações de parentesco das fêmeas juvenis não serem conhecidas, não podemos afirmar até que ponto o apoio concedido, pelo macho e fêmea dominante, não representa uma protecção parental. Por outro lado, dada a intervenção sempre a favor do agressor, numa hierarquia linear, e como tal com risco reduzido para o apoiante, a participação das fêmeas juvenis aponta para mecanismos de aquisição de estatuto. A sua ligação ao macho e fêmea dominante ajudam a explicar a sua posição hierárquica elevada (logo abaixo da fêmea dominante) na hierarquia de fêmeas. O papel interventivo dos indivíduos mais dominantes na formação de alianças, independentemente da catagem recebida, foi verificado anteriormente por Hemelrijk (1990), tendo este sugerido que as intervenções dos indivíduos dominantes servem sobretudo para a manutenção da estrutura hierárquica do grupo.
Os dados do presente estudo referem-se a um curto período de tempo, pelo que poderíamos ainda invocar algum tipo de mecanismo de investimentos nas relações a longo prazo. No entanto, evidências que os primatas não-humanos possuam as capacidades cognitivas que lhes permitam planear esses objectivos, não foram ainda claramente demonstradas. Na falta de outras opções, considerar que os indivíduos de baixo estatuto são mais dependentes dos laços sociais, e como tal, investem mais na sua formação, parece ser um raciocínio algo circular, especialmente na ausência de uma compreensão exacta do que significa dependência de laços sociais.
A inexistência de uma relação entre a catagem e a formação de alianças sugere que outros bens podem estar a ser trocados por esta. Um exemplo, poderia ser a tolerância nos locais de alimentação. Apesar de ter sido observado que os indivíduos dominantes restringiam o acesso inicial dos subordinados aos tabuleiros de comida, não existiam sinais exteriores de que os indivíduos subordinados apresentavam problemas visíveis de subnutrição. Podemos considerar ainda a catagem, como mecanismo de evitamento de potenciais conflitos (Aureli et al., 1995; Judge & de Waal, 1997), dada as condições restritas de cativeiro e o papel da catagem como regulador da tensão (Boccia, 1987).
A perspectiva de mercado aplicada à catagem baseia-se na ideia de que a troca é um elemento essencial em muitas interacções de benefício mútuo (Noë & Hammerstein, 1995). Embora a teoria dos mercados biológicos ajude a explicar estas interacções, muitos aspectos destas transacções sociais precisam ainda de ser clarificados.
REFERÊNCIAS
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227-267. [ Links ]
Barrett, L., & Henzi, S. P. (2002). Constraints on relationship formation among female primates. Behaviour, 139, 263-289.
Barrett, L., Gaynor, D., & Henzi, S. P. (2002). A dynamic interaction between aggression and grooming reciprocity among female chacma baboons. Animal Behaviour, 63, 1047-1053.
Barrett, L., Henzi, S. P., Weingrill, T., Lycett, J. E., & Hill, R. A. (1999). Market forces predict grooming reciprocity in female baboons ( Papio hamdryas cynocephalus). American Journal of Primatology, 47, 321-334.
Bentley-Condit, V. K., & Smith, E. (1999). Female dominance and female social relationships among yellow baboons (Papio hamadryas cynocephalus). American Journal of Primatology, 47, 321-334.
di Bitetti, M. S. (1997). Evidence for an important social role of allogrooming in a platyrrhine primate. Animal Behaviour, 54, 199-211.
Chance, M. R. A. (1967). Attention structure as the basis of primate rank o rders. Man, 2, 503-518.
Cooper, M. A., & Bernstein, I. S. (2000). Social grooming in assamese macaques (Macaca assamensis). American Journal of Primatology, 50, 77-85.
Corradino, C. (1990). Proximity structure in a captive colony of Japanese monkeys (Macaca fuscata fuscata): An application of multidimensional scalling. Primates, 31, 351-362.
Dunbar, R. I. M. (1991). Functional significance of social grooming in primates. Folia Primatologica, 57, 121-131.
Fairbanks, L. (1976). A comparative analyses of subgroup structure and spatial relationships in captive baboons and squirrel monkeys. Primates, 17, 291-300.
Fairbanks, L. (1980). Relationships among adult females in captive vervet monkeys: Testing a model of rank-related attractiveness. Animal Behaviour, 28, 853-859.
Franz, C. (1999). Allogrooming behaviour and grooming site preferences in captive bonobos (Pan paniscus): Association with female dominance. American Journal of Primatology, 20, 525-546.
Gust, D. A. (1995). Moving up the dominance hierarchy in young sooty mangabeys. Animal Behaviour, 50, 15-21.
Hector, A. C. K., Seyfarth, R. M., & Raleigh, M. J. (1989). Male parental care, female choice and the effect of an audience in vervet monkeys. Animal Behaviour, 38, 262-271.
Hemelrijk, C. K. (1990). A matrix partial correlation test used in investigations of reciprocity and other social interaction patterns at group level. Journal of Theoretical Biology, 143, 405-420.
Hemelrijk, C. K. (1994). Support for being groomed in long-tailed macaques, Macaca fascicularis. Animal Behaviour, 48, 479-481.
Hemelrijk, C. K., & Ek, A. (1991). Reciprocity and interchange of grooming and ‘support’ in captive chimpanzees. Animal Behaviour, 41, 923-935.
Henzi, S. P., & Barrett, L. (1999). The value of grooming to female primates. Primates, 40, 47-59.
Henzi, S. P., & Barrett, L. (2002). Infants as a commodity in a baboon market. Animal Behaviour, 63, 915-921.
Lazaro-Perea, C., Arruda, M. D. F., & Snowdown, C. T. (2004). Grooming as a reward? Social function of grooming between females in cooperatively breeding marmosets. Animal Behaviour, 67, 627-636.
Leinfelder, I., de Vries, H., Deleu, R., & Nelissen, M. (2001). Rank and grooming reciprocity among females in a mixed-sex group of captive hamadryas baboons. American Journal of Primatology, 55, 25-42.
Manson, J. H., Rose, L. M., Perry, S., & Gros-Louis, J. (1999). Dynamics of female-female relationships in wild Cebus capucinus: Data from two Costa Rican sites. International Journal of Primatology, 20, 679-706.
Matheson, M. D., & Bernstein, I. S. (2000). Grooming, social bonding, and agonistic aiding in rhesus monkeys. American Journal of Primatology, 51, 177-186.
McGuire, M. T. (1974). St. Kitts vervets. Contributions to Primatology (vol. 1). Basel: S. Karger AG.
McNelis, N. L., & Boatright-Horowitz, S. L. (1998). Social monitoring in a primate group: The relationship between visual attention and hierarchical ranks. Animal Cognition, 1, 65-69.
Melnick, D. J., & Pearl, M. C. (1987). Cercopithecines in multimale groups: Genetic diversity and population structure. In B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, & T. T. Struhsaker (Eds.), Primate Societies (pp. 121-134). Chicago: The University of Chicago Press.
Muroyama, Y. (1991). Mutual reciprocity of grooming in female Japanese macaques (Macaca fuscata). Behaviour, 119, 161-170.
Muroyama, Y. (1994). Exchange of grooming for allomothering in female patas monkeys. Behaviour, 128, 103-119.
Muroyama, Y. (1996). Decision making in grooming by Japanese macaques (Macaca fuscata). International Journal of Primatology, 17, 817-830.
Nakamichi, M. (2003). Age related differences in social grooming among adult female Japanese monkeys (Macaca fuscata). Primates, 44, 239-246.
Nakamichi, M., & Schizawa, Y. (2003). Distribution of grooming among adult females in a large, free-ranging group of Japanes macaques. International Journal of Primatology, 24, 607-625.
Noë, R., & Hammerstein, P. (1994). Biological markets: Supply and demand determine the effect of partner choice in cooperation, mutualism and mating. Behavioral Ecology and Sociobiology, 35, 1-11.
Noë, R., & Hammerstein, P. (1995). Biological markets.Trends in Ecology and Evolution, 10, 336-339.
Parr, L. A., Matheson, M. D., Bernstein, I. S., & de Waal, F. B. M. (1997). Grooming down the hierarchy: Allogrooming in captive brown capuchin monkeys, Cebus apella. Animal Behaviour, 54, 361-367.
Pastor-Nieto, R. (2001). Grooming, kinship, and co-feeding in captive spider monkeys (Ateles geoffroyi). Zoo Biology, 20, 293-303.
Payne, H. F. P., Lawes, M. J., & Henzi, S. P. (2003). Competition and the exchange of grooming among female samango monkeys (Cercopithecus mitis erythrarchus). Behaviour, 140, 453-471.
Raleigh, M. J., & McGuire, M. T. (1989). Female influences on male dominance acquisition in captive vervet monkeys, Cercopithecus aethiops sabaeus. Animal Behaviour, 38, 59-67.
Rowell, T. E. (1971). Organization of caged groups of Cercopithecus monkeys. Animal Behaviour, 16, 625-645.
Sambrook, T. D., Whiten, A., & Strum, S. C. (1995). Priority of access and grooming patterns of females in a large and a small group of olive baboons. Animal Behaviour, 50, 1667-1682.
Schino, G. (2001). Grooming, competition and social rank among female primates: A meta-analysis. Animal Behaviour, 62, 265-271.
Schino, G., Ventura R., & Troisi, A. (2003). Grooming among female Japanese macaques: distinguishing between reciprocation and interchange. Behavioral Ecology, 14, 887-891.
Seyfarth, R. M. (1977). A model of social grooming among adult female monkeys. Journal of Theoretical Biology, 65, 671-698.
Seyfarth, R. M. (1980). The distribution of grooming and related behaviours among adult female vervet monkeys. Animal Behaviour, 28, 798-813.
Seyfarth, R. M., & Cheney, D. L. (1984). Grooming, alliances and reciprocal altruism in vervet monkeys. Nature, 308, 541-543.
Silk, J. B. (1999). Why are infants so attractive to others? The form a nd function of infant handling in bonnet macaques. Animal Behaviour, 57, 1021-1032.
Silk, J. B. (2002a). Kin selection in primate groups. International Journal of Primatology, 23, 849-875.
Silk, J. B. (2002b). Using the ‘F’-word in primatology. Behaviour, 139, 421-446.
Silk, J. B., Seyfarth, R. M., & Cheney, D. L. (1999). The structure of social relationships among female savana baboons in Moremi reserve, Botswana. Behaviour, 136, 679-703.
Silk, J. B., Seyfarth, R. M., & Altmann, J. (2004). Patterns of coalition formation by adult female baboons in Amboseli, Kenya. Animal Behaviour, 67, 573-582.
Strayer, F. F., & Gariepy, J. L. (1986). The structure of social attention and its coordination with coesive and dispersive activities in captive groups of squirrel monkeys. In D. M. Taub, & F. A. King (Eds.), Current perspectives in primate social dynamics (pp. 99-111). New York: Van Nostrand Reinhold.
de Waal, F. B. M. (1991). Rank distance as central feature of rhesus monkey social organization: a sociometric analysis. Animal Behaviour, 41, 383-395.
de Waal, F. B. M. (1997). The chimpanzee’s service economy: Food for grooming. Evolution and Human Behavior, 18, 375-386.
de Waal, F. B. M., & Lutrell, L. M. (1986). The similarity principle underlying social bonding among female rhesus monkeys. Folia Primatologica, 46, 215-234.
Watts, D. P. (2000). Grooming between male chimpanzees at Ngogo, Kibale National Park. II. Influence of male rank and possible competition for partners. International Journal of Primatology, 21, 211-238.
White, F. J., & Chapman, C. A. (1994). Contrasting chimpanzees and bonobos: Nearest neighbour distances and choices. Folia Primatologica, 63, 181-191.
(*) Bolseiro FCT (FCT-SFRH/BD/27489/2006), UIPCDE, Instituto Superior de Psicologia Aplicada, Lisboa. E-mail: joaordaniel@gmail.com
(**) UIPCDE, Instituto Superior de Psicologia Aplicada, Lisboa.
(***) Faculdade de Ciências da Universidade de Lisboa, Centro de Biologia Ambiental.

