










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkSilva Lusitana
versão impressa ISSN 0870-6352
Silva Lus. vol.22 no.1 Lisboa jun. 2014
ARTIGOS
Competição em Nível de Árvore Individual em uma Floresta Estacional Semidecidual
Competition indices in individual tree level in a Semideciduous Montana forest
La concurrence sur chaque arbre de la Forêt Semi-Décidue saisonnière
*Renato Castro, **Carlos Soares, **Helio Leite, **Agostinho Souza, ***Fabrina Martins, ****Gilciano Nogueira, ****Márcio Oliveira e Fabrício Silva1
*Doutor em Ciência Florestal, Universidade Federal de São João del Rey, Campus Sete Lagoas, Departamento de Ciências Agrárias. CEP 70904-970 Sete Lagoas, MG, BRASIL. E-mail: castrorvo@ymail.com
**Doutor em Ciência Florestal, 1Estudante de Mestrado em Ciência Florestal, Universidade Federal de Viçosa, Departamento de Engenharia Florestal. Avenida Peter Henry Rolfs, s/n, Campus Universitário, CEP 36570-000 Viçosa, MG BRASIL. E-mails: csoares@ufv.br; hgleite@gmail.com; alsouza@ufv.br; fabricioardufv@gmail.com
***Doutora em Ciência Florestal, Universidade Federal de Itajubá, Instituto de Recursos Naturais. Av. BPS nº 1.303, Bairro Pinheirinho, CEP 37500-903 Itajubá, MG, BRASIL. E-mail: fabrinabm@gmail.com
****Doutor em Ciência Florestal, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Departamento de Engenharia Florestal. Rua da Glória nº 187, Centro, CEP 39100-000 Diamantina, MG, BRASIL. E-mails: nogueirags@gmail.com; marcioromarco@yahoo.com.br
RESUMO
Este estudo teve como objetivo avaliar a correlação entre diferentes índices de competição com o crescimento diamétrico (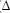 dap), crescimento em altura (Ht) e probabilidade de mortalidade (Pm) em árvores individuais de um fragmento de Floresta Estacional Semidecidual Montana em estágio médio de sucessão ecológica. Foram avaliadas três categorias de índices, sendo cinco índices independentes (IID), dois dependentes (IDD) e quatro índices semi-independentes da distância (ISD). Para os índices independentes e semi-independentes da distância foram avaliados dois raios de vizinhança: seis e três metros. Os dados empregados foram provenientes de dez parcelas monitoradas durante 14 anos, em cinco ocasiões, localizadas no município de Viçosa, Minas Gerais, Brasil. Os índices de competição foram avaliados a partir de análises gráficas e coeficientes de correlações lineares entre cada índice e as variáveis dap, Ht e Pm. As correlações foram calculadas para todas as espécies em conjunto e para 36 espécies isoladamente, que apresentaram densidade (número de árvores) maior ou igual a 10 indivíduos. De maneira geral, todas as categorias de índices de competição (IID, IDD e ISD) apresentaram índices correlacionados com o crescimento e mortalidade de árvores individuais, com superioridade dos índices semi-independentes da distância. Em relação ao raio de competição, para os índices dependentes e semi-independentes, foi observado superioridade para o raio de 6 m, para todas as variáveis. Assim sendo, observou-se a potencialidade de todas as categorias de índices de competição em estudos de crescimento e produção em nível de árvore individual em Floresta Estacional Semidecidual Montana, em nível médio de sucessão ecológica. Os resultados servirão de subsídio para a inclusão de índices de competição em modelos de crescimento e produção em nível de árvore individual em florestas inequiâneas, especialmente em Floresta Estacional Semidecidual Montana, em nível médio de sucessão ecológica.
dap), crescimento em altura (Ht) e probabilidade de mortalidade (Pm) em árvores individuais de um fragmento de Floresta Estacional Semidecidual Montana em estágio médio de sucessão ecológica. Foram avaliadas três categorias de índices, sendo cinco índices independentes (IID), dois dependentes (IDD) e quatro índices semi-independentes da distância (ISD). Para os índices independentes e semi-independentes da distância foram avaliados dois raios de vizinhança: seis e três metros. Os dados empregados foram provenientes de dez parcelas monitoradas durante 14 anos, em cinco ocasiões, localizadas no município de Viçosa, Minas Gerais, Brasil. Os índices de competição foram avaliados a partir de análises gráficas e coeficientes de correlações lineares entre cada índice e as variáveis dap, Ht e Pm. As correlações foram calculadas para todas as espécies em conjunto e para 36 espécies isoladamente, que apresentaram densidade (número de árvores) maior ou igual a 10 indivíduos. De maneira geral, todas as categorias de índices de competição (IID, IDD e ISD) apresentaram índices correlacionados com o crescimento e mortalidade de árvores individuais, com superioridade dos índices semi-independentes da distância. Em relação ao raio de competição, para os índices dependentes e semi-independentes, foi observado superioridade para o raio de 6 m, para todas as variáveis. Assim sendo, observou-se a potencialidade de todas as categorias de índices de competição em estudos de crescimento e produção em nível de árvore individual em Floresta Estacional Semidecidual Montana, em nível médio de sucessão ecológica. Os resultados servirão de subsídio para a inclusão de índices de competição em modelos de crescimento e produção em nível de árvore individual em florestas inequiâneas, especialmente em Floresta Estacional Semidecidual Montana, em nível médio de sucessão ecológica.
Palavras-chave: Floresta Estacional Semidecidual Montana, índices dependentes da distância, índices independentes da distância, índices semi-independentes da distância, correlação linear
ABSTRACT
This study aimed to evaluate the correlation between different levels of competition with the diameter growth (dap), height growth (Ht) and probability of mortality (Pm) of individual trees in a Semideciduous forest fragment in Montana middle stage of ecological succession. We evaluated three classes of indicators, five independent indices (IID), two dependent (IDD) and four semi-independent indices of distance (ISD). To indexes independent and semi-independent of the distance were assessed two neighborhood rays: six and three meters. The data used were from ten plots monitored for 14 years, on five occasions, located in Viçosa, Minas Gerais, Brazil. The competition indexes were evaluated from graphical analysis and linear correlation coefficients between each index and the variables dap, Ht and Pm. Correlations were calculated for all species together and for 36 species alone, which had density greater than or equal to 10 individuals. In general, all categories of competition indexes (IID, IDD and ISD) had indices correlated with the growth and mortality of individual trees, with higher rates of indices semi-independent of the distance. In relation to the radius of competition for the contents semi-dependent and independent superiority was observed for the radius of 6 m, for all variables. Thus, we noticed the potential for all categories of competition indexes in studies of growth and production level of individual tree in Semideciduous Montana forest in average level of ecological succession. The results will inform the inclusion of competition indexes in models of growth and production in individual tree basis in uneven-aged forests, especially Semideciduous Montana forest in average level of ecological succession.
Key words: Semideciduous Montana forest, distance-dependent indices, indices independent of distance, indices semi-independent of distance, linear correlation
RÉSUMÉ
Cette étude visait à évaluer la corrélation entre les différents niveaux de compétition avec la croissance en diamètre (dap), la croissance en hauteur (Ht) et la probabilité de mortalité (Pm) dans les arbres individuels d'un fragment de forêt semi-décidue saisonnière dans le Montana, étape intermédiaire de la succession écologique. Trois catégories d'indices, cinq indices indépendants (IID), deux dépendant (IDD) et quatre indices semi- indépendants de la distance (ISD) ont été évalués. Pour les indices indépendants et semi- indépendants de la distance, deux rayons ont été évalués: six et trois mètres: Les données utilisées provenaient de dix parcelles suivies pendant 14 ans à cinq reprises, situés dans Viçosa, Minas Gerais, Brésil. Les indices de compétition ont été évalués à partir de l'analyse graphique et les coefficients de corrélation linéaire entre chaque indice et les variables dap, Ht et Pm. Les corrélations ont été calculés pour toutes les espèces ensemble et 36 espèces, individuelles, qui ont montré une densité supérieure ou égale à 10 individus. En général, toutes les catégories d'indices de la concurrence (IID, IDD et ISD) ont été corrélées avec la croissance et la mortalité des arbres individuels, avec une supériorité des indices semi-indépendants des indices de distance. En ce qui concerne le rayon de la concurrence, pour les indices dépendants et semi-indépendants une supériorité par rapport au rayon de 6 m a été observée pour toutes les variables. Ainsi, le potentiel a été observé pour toutes les catégories d'indices de la concurrence dans les études de la croissance et de la production pour l'arbre individuel en forêt semi-décidue dans le Montana, en niveau moyen de la succession écologique. Les résultats serviront à informer l'inclusion des indices de la concurrence dans les modèles de croissance et de production pour l'arbre individuel dans les forêts, en particulier dans la forêt semi-décidue Montana en niveau moyen de la succession écologique.
Mots-clés: Forêt Semi-Décidue Saisonnière Montana, index dépendant de la distance, indépendamment des rapports de distance, indices semi-indépendants de la distance, corrélation linéaire
Introdução
Índices de competição são importantes para a modelagem de crescimento e produção em florestas tropicais (Foli, 1993). O emprego desses índices permite a determinação do nível de supressão de cada árvore em relação às suas competidoras (Martins et al., 2011).
Eles estão entre as variáveis explicativas mais utilizadas para se predizer o crescimento e a mortalidade de árvores individuais (Radtke et al., 2003, Peltoniemi e Mäkipää, 2011). O efeito negativo da competição sobre o crescimento e mortalidade das árvores é confirmado por diversos autores (Wykoff, 1990; Quicke et al., 1994; BigGing e Dobbertin, 1995; Hasenauer e Monserud, 1996; Monserud e Sterba, 1996; Sterba et al., 2002). Porém, a avaliação da competição constitui um problema contínuo na área florestal, devido principalmente à dificuldade de sua quantificação (Holmes e Reed, 1991; Oheimb et al., 2011).
Muitos estudos sobre quantificação da competição entre as árvores foram conduzidos em florestas de uma ou poucas espécies (Glover e Hool, 1979; Martin e Ek, 1984; BigGing e Dobbertin, 1992, 1995; Pretzsch, 2002; Martins et al., 2011; Castro, 2011), sendo escassos os estudos em áreas com elevada riqueza de espécies (Hamilton, 1986; Zhao et al., 2006; Oheimb et al., 2011).
Na literatura são conhecidos os índices de competição dependentes da distância (IDD), independentes da distância (IID) e semi-independentes da distância (ISD) (Alig et al., 1984; Stage e Ledermann, 2008; ContReras et al., 2011). Os índices de competição diferem conceitualmente em função do critério utilizado para expressar as relações entre as árvores.
Nos índices de competição dependentes da distância são atribuídas coordenadas espaciais para cada árvore, as quais permitem ponderar a competição entre cada árvore e as circunvizinhas pela distância, sendo mais complexos e exigentes em termos de coleta de dados (Hynynen, 1995). Uma grande variedade destes índices foi introduzida em estudos de crescimento e produção de árvores em florestas homogêneas na América do Norte (BiGging e Dobbertin, 1992) e Europa (Tomé e Burkhart, 1989).
Os índices independentes da distância não utilizam informações da distribuição espacial dos indivíduos, sendo quantificados pela razão entre diâmetro e/ou altura das árvores e variáveis do povoamento, tais como área basal, diâmetro médio e altura média (Daniels et al., 1986; Prodan et al., 1997; Della-Flora, 2001). Os vizinhos (árvores competidoras) da árvore-objeto (aquela em estudo) são definidos pelas árvores incluídas na mesma unidade amostral (Sironen et al., 2002).
Já para o cálculo dos índices de competição semi-independentes da distância, são aplicados os índices independentes da distância, entretanto, espacialmente restritos a um menor conjunto de árvores vizinhas (aquelas mais próximas), ao invés de todas as árvores da unidade amostral (Stage e Ledermann, 2008; Lederman, 2010; Contreras et al., 2011).
A eficiência da modelagem do crescimento e da mortalidade em nível de árvore individual pode ser maior com o uso de índices de competição (Lorimer, 1983; Castro, 2011). Martin e Ek (1984), ao incluírem índices de competição independentes da distância nos modelos de crescimento de Pinus resinosa, obtiveram resultados superiores em relação aos modelos sem a utilização de índices. PUKKALA e Kolstrem (1987) avaliaram vários índices dependentes da distância e verificaram que os melhores índices poderiam explicar até 50% da variação de crescimento em diâmetro para Pinus sylvestris. Biging e Dobbertin (1995), ao revisarem vários estudos em relação aos ganhos no poder de predição com a utilização de índices de competição, concluíram que a superioridade de um índice é verificada somente para determinadas situações e espécies.
Na avaliação do desempenho dos índices de competição é frequente a estimação da correlação com as variáveis de interesse, como o crescimento em diâmetro, a área basal e a altura (Daniels et al., 1986; Weber et al., 2008) e a probabilidade de mortalidade (Martins et al., 2011). Em florestas naturais tropicais ainda são escassos trabalhos avaliando índices de competição e a sua relação com o crescimento das árvores.
O presente estudo teve como objetivo estimar e avaliar índices de competição independentes, dependentes e semi-independentes da distância em árvores individuais de um fragmento de Floresta Estacional Semidecidual Montana, e identificar aqueles índices que mais se correlacionam com o crescimento diamétrico, crescimento em altura e probabilidade de mortalidade de árvores.
Material e métodos
Dados
Este estudo foi realizado no município de Viçosa, Minas Gerais, Brasil, em um fragmento florestal pertencente à Universidade Federal de Viçosa, que possui 17 ha de área (42°52'W e 42° 50'W de longitude e 20°44'S e 20°47'S de latitude).
O clima local, segundo a classificação de Köppen, é do tipo Cwa, temperado quente, com média das temperaturas máximas de 26°C e média das temperaturas mínima de 14°C, caracterizado por verões quentes e chuvosos e invernos frios e secos (Amaro, 2010).
A precipitação média anual durante o período desse estudo foi de 1.280 mm (Inmet, 2012). O solo apresenta a predominância de duas classes: no topo e nas encostas das elevações ocorrem os latossolos vermelho-amarelo álico, enquanto nos terraços o solo predominante é o Podzólico Vermelho-Amarelo câmbico (Rezende, 1971). A topografia da região é acidentada, com vales estreitos e úmidos e altitude variando de 600 a 970 metros (Corrêa, 1984).
O município de Viçosa pertence à região fitoecológica de Floresta Estacional Semidecidual Montana (Veloso et al., 1991) e o fragmento estudado encontra-se em estágio médio de sucessão (Figueiredo, 2011).
Para o presente trabalho, foi amostrada uma área de um hectare, constituída por dez parcelas retangulares, não contínuas, de 1.000 m² cada (20m x 50m), distribuídas aleatoriamente no fragmento (Meira Neto e Martins, 2000).
Em cada parcela, as árvores foram identificadas e mensurados os diâmetros a 1,3 m de altura (dap) e altura total (Ht) de todas as árvores com dap = 5 cm, nos anos de 1994, 1997, 2000, 2004 e 2008. Para cada árvore foram determinadas as coordenadas x e y em relação à origem da sua parcela e criado um mapa de localização dos indivíduos com auxílio do software ArcGis 10. Para o cálculo das distâncias entre cada árvore e as suas competidoras, foi utilizada a ferramenta Point Distance disponível no software ArcGis 10.
A identificação de todo o material botânico coletado foi feita por meio de comparações com o material do Herbário do Departamento de Biologia Vegetal da Universidade Federal de Viçosa (VIC-UFV) e consultas a especialistas. O sistema de classificação adotado foi o APG III (2009). Os indivíduos foram classificados em grupos ecológicos segundo Gandolfi et al. (1995).
As principais características quantitativas do povoamento estudado são apresentadas no Quadro 1.
Índices de competição
Para cada fuste, em cada medição, foram estimados índices de competição independentes da distância (IID), dependentes da distância (IDD) e semi-independentes da distância (ISD). Para automatizar o cálculo dos índices foi desenvolvido um aplicativo para cada categoria de índice em ambiente Visual Basic for Applications, utilizando o Microsoft Excel ®, versão 2007.
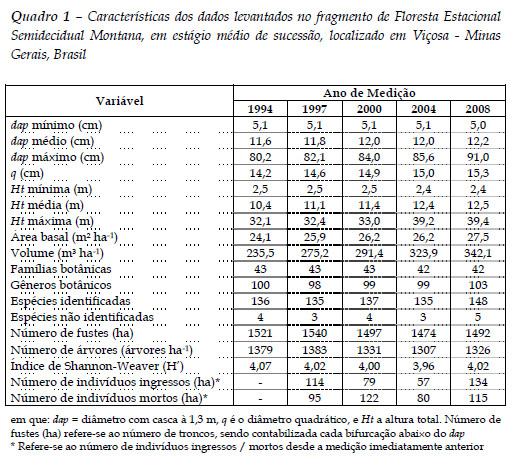
Para os índices independentes e semi-independentes da distância, foram utilizados dois raios de vizinhança, de seis e três metros, aos quais correspondem, em média, a 16 vizinhos para o raio competitivo de seis metros, e 3,3 vizinhos no raio de três metros, conforme apresentado na Figura 1. Os vizinhos só foram qualificados como competidores se tivessem o nível de inclusão mínimo de dap= 5cm.
Não foram avaliados raios de competição maiores devido à limitação de tamanho das unidades amostrais (parcelas), conforme já reportado por Biging e Dobbertin (1995). Em raios maiores, as árvores próximas à bordadura das parcelas seriam sensivelmente influenciadas no cálculo dos índices, visto que menos competidores são observados, o que resulta em uma subestimação da competição.
Hegyi (1974) determinou o número de competidores utilizando um raio de 3,05 m a partir da árvore-objeto e Béland et al. (2003), variou o raio de competição entre 3 e 9 m, e indicou o raio competitivo de 6 m após a simulação em modelos de regressão, ambos em povoamentos de Pinus.
A relação funcional de cada índice de competição utilizado é apresentada no Quadro 2.
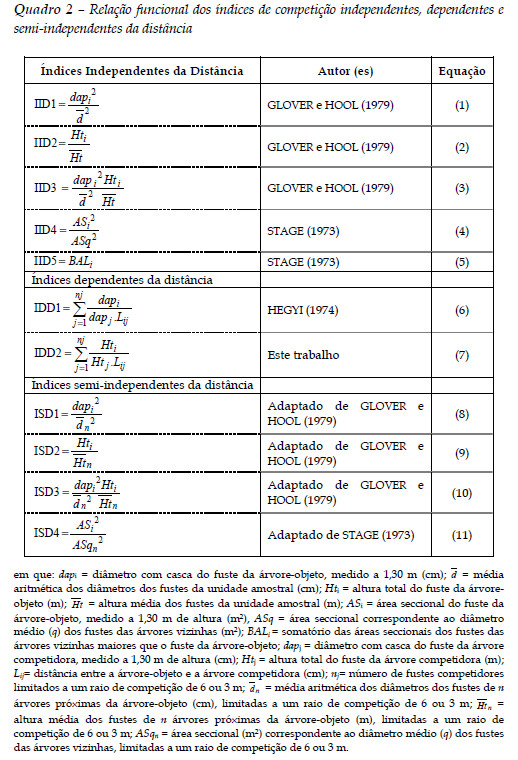
em que: dapi = diâmetro com casca do fuste da árvore-objeto, medido a 1,30 m (cm); = média aritmética dos diâmetros dos fustes da unidade amostral (cm); Hti = altura total do fuste da árvore-objeto (m); = altura média dos fustes da unidade amostral (m); ASi = área seccional do fuste da árvore-objeto, medido a 1,30 m de altura (m²), ASq = área seccional correspondente ao diâmetro médio (q) dos fustes das árvores vizinhas (m²); BALi = somatório das áreas seccionais dos fustes das árvores vizinhas maiores que o fuste da árvore-objeto; dapj = diâmetro com casca do fuste da árvore competidora, medido a 1,30 m de altura (cm); Htj = altura total do fuste da árvore competidora (m); Lij= distância entre a árvore-objeto e a árvore competidora (cm); nj= número de fustes competidores limitados a um raio de competição de 6 ou 3 m; = média aritmética dos diâmetros dos fustes de n árvores próximas da árvore-objeto (cm), limitadas a um raio de competição de 6 ou 3 m; = altura média dos fustes de n árvores próximas da árvore-objeto (m), limitadas a um raio de competição de 6 ou 3 m; ASqn = área seccional (m²) correspondente ao diâmetro médio (q) dos fustes das árvores vizinhas, limitadas a um raio de competição de 6 ou 3 m.
Avaliação dos índices de competição
Depois de calculados os índices de competição, foram realizadas análises gráficas relacionando o crescimento anual em diâmetro e em altura com cada índice estudado. Em seguida foi elaborada uma matriz de correlação linear entre os índices e o crescimento anual em diâmetro (dap), crescimento anual em altura (Ht), e a probabilidade de mortalidade (Pm), obtidos entre cada intervalo de medição. As significâncias das correlações foram avaliadas ao nível de 95% de probabilidade.
Foram estimadas correlações considerando todas as espécies em conjunto e também individualmente para 36 espécies, selecionadas por apresentarem densidade maior ou igual a 10 indivíduos.
A probabilidade de mortalidade (Pm) foi obtida calculando-se a proporção de fustes mortos por classe de diâmetro para cada parcela, entre os intervalos de medição, por meio da seguinte expressão (adaptado de Martins et al., 2011; Castro, 2011):

em que nj1 é o número de fustes vivos na j-ésima classe de diâmetro, no início do período, nj2 é o número de fustes vivos na j-ésima classe de diâmetro no fim do período, descontados os indivíduos ingressos entre os períodos sequentes (Ij1j2).
Utilizando as correlações estimadas para cada uma das 36 espécies, foi calculado um valor de superioridade (VS) para cada índice de competição, em relação a cada variável estudada (dap, Ht e Pm). O valor de superioridade para cada índice i foi calculado pelo somatório do valor de importância das espécies que apresentaram esse índice i com o maior coeficiente de correlação significativa (em módulo), de acordo com a seguinte fórmula, desenvolvida para este estudo:

em que VSij é valor de superioridade do índice i, relativo à categoria de índices j (IID, IDD ou ISD); IVI é o índice de valor de importância da espécie k, caso a correlação seja significativa; w é o número de índices existentes na categoria j; y é o número total de índices, considerando todas as categorias. O valor de w utilizado no cálculo do VS de cada índice foi igual a 5, 4 e 8, respetivamente para as categorias dos índices IID, IDD e ISD e o valor de y foi igual a 17.
Dentro de cada categoria de índices e cada variável, aquele índice de competição que apresentou maior valor de superioridade (VS) foi considerado como sendo o mais indicado para ser utilizado em modelos para todas as espécies em conjunto. O termo w da equação 13 foi utilizado como ponderação entre índices de diferentes categorias (que apresentam diferentes quantidades de índices). Assim foi possível a comparação da superioridade entre índices de diferentes categorias.
Resultados e discussão
As tendências médias dos índices de competição apresentaram coerência em relação ao crescimento em diâmetro e em altura das árvores (Figura 2).
Para os índices IID1 a IID4 e de ISD1 a ISD4 (eq. 1 a 4 e 8 a 11) foi observado que quanto menor o crescimento em diâmetro e altura, menor o índice de competição. Estes índices são derivados da razão entre uma variável dendrométrica ou uma combinação de variáveis dendrométricas do fuste da árvore-objeto em relação à média das mesmas variáveis dos fustes das árvores qualificadas como vizinhas. Isto indica que, de forma geral, fustes que apresentam sua variável dendrométrica inferior à média dos fustes vizinhos apresentarão menores índices, resultando em menor crescimento. Essa mesma tendência foi observada por outros autores como Tomé e Burkhart (1989) e Martins et al. (2011).
O índice IID5 apresenta menores valores para os fustes com maior crescimento em diâmetro e altura, contrariamente ao que ocorre com os índices anteriores. Tal fato foi explicado por Martins et al. (2011), pois consiste no somatório das áreas seccionais dos fustes maiores que o fuste da árvore-objeto, de forma que os menores fustes apresentam maiores valores no referido índice. Quanto maior o IID5, maior o nível de competição.
Os índices dependentes da distância (eq. 6 e 7), com os raios competitivos de 6 e 3 m, também apresentaram tendência de aumento à medida que são observados maiores valores de crescimento, por também serem quantificados a partir de uma razão entre variáveis dendrométricas. Entretanto, como são ponderados pela distância entre as vizinhas e a árvore-objeto, o comportamento dos índices para fustes de mesmo tamanho pode ser variado, devido à distribuição das árvores presentes nas parcelas.
Apesar da coerência apresentada pelos índices de competição (Figura 2), quando realizadas as análises de correlações lineares com o crescimento em diâmetro, o crescimento em altura e a probabilidade de mortalidade para todas as espécies em conjunto, foram observadas correlações inferiores a 0,30, entretanto, em sua maioria, significativas a 95% de probabilidade (Quadro 3).
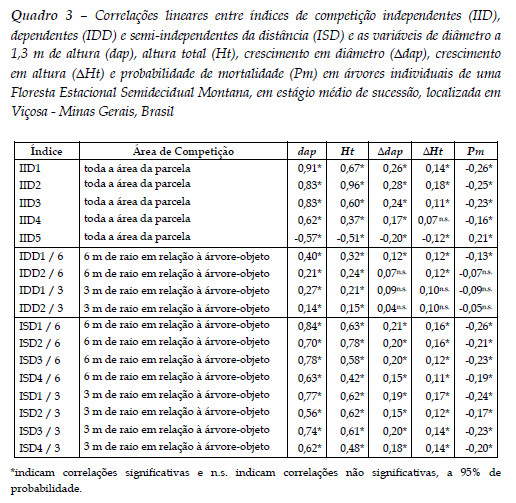
As baixas correlações são atribuídas à elevada heterogeneidade de espécies na floresta, idades das árvores e condições gerais de crescimento e sanidade, dentre outros.
Os índices de competição são calculados em função dos tamanhos dos fustes das árvores e, para alguns deles, utilizando a informação de distância. Dessa forma, em fustes de mesmo porte são atribuídos índices de competição com valores muito próximos, mesmo que não possuam as mesmas taxas de crescimento.
Em florestas tropicais, existem grandes variações no crescimento (diâmetro e altura) em indivíduos de mesmo tamanho. Enquanto algumas espécies crescem muito em um período, outras não crescem. Além disso, neste estudo não foram avaliados atributos de tamanho das copas das árvores para quantificação dos índices, os quais poderiam colaborar com o entendimento das variáveis, como reportado por Canham et al. (2004) e Coates et al. (2009).
Contrariamente, em florestas equiâneas, árvores de mesmo tamanho possuem taxas de crescimento semelhantes. Assim, podem ser observados valores de correlações superiores, conforme apresentado por Martins et al. (2011).
Os índices apresentaram sinais coerentes de correlação com a probabilidade de mortalidade (Quadro 3). Com o aumento da probabilidade de mortalidade houve tendência de aumento do índice IID5 (correlação positiva) e tendência de diminuição dos demais índices (correlações negativas). Isso indica que, em média, árvores menores possuem maiores probabilidades de morrer. Rossi et al., (2007) também afirma que, em florestas inequiâneas, é esperado uma maior probabilidade de mortalidade em árvores das classes de diâmetro inferiores, visto que representam as árvores suprimidas da floresta.
Ainda não foram desenvolvidos índices de competição que relacionam conceitos de competição intra-específica (entre indivíduos da mesma espécie) e interespecífica (entre indivíduos de espécies diferentes), que poderia aumentar sobremaneira o entendimento da competição entre árvores em florestas inequiâneas.
As correlações lineares entre os índices de competição independentes, dependentes e semi-independentes da distância e as variáveis dap, Ht e Pm por espécie, para aquelas que apresentaram densidade maior ou igual a 10 indivíduos, são apresentadas no Quadro 4, sendo apresentado apenas aquele índice (dentre os 17 índices[1]), com a maior correlação (em módulo) de cada categoria e para cada espécie.
As correlações significativas para o crescimento em diâmetro variaram entre -0,12 e -0,55, e 0,17 e 0,58. As maiores correlações positivas foram observadas para as espécies Tapirira guianensis (ISD4/3), Platypodium elegans (IID4) e Newtonia contorta (IID2), com correlações variando entre 0,54 e 0,58. As maiores correlações negativas, em módulo, foram observadas para as espécies Endlicheria paniculata (ISD2/6) e Allophylus edulis (IDD2/3), com correlações de -0,42 e -0,55.
Para o crescimento em altura, foram constatadas correlações significativas variando entre -0,18 e -0,42, e 0,15 e 0,54. Maiores correlações positivas foram verificadas para as espécies Dalbergia nigra (IDD2/6), Luehea speciosa (IID4) e Mabea fistullifera (IDD2/6), variando entre 0,51 a 0,54, e as correlações negativas para as espécies Endlicheria paniculata (IID2) e Casearia arborea (ISD2/3), iguais a -0,32 e -0,42, respetivamente.
Ganhos consideráveis nos valores de correlações, quando calculadas em nível de espécie, foram observados para a variável probabilidade de mortalidade. As correlações significativas variaram entre -0,16 e -0,49, e 0,24 e 0,55. As maiores correlações positivas foram observadas para as espécies Piptadenia gonoacantha (IID5), Prunus sellowii (IID5) e Maytenus robusta (IID4), variando entre 0,48 e 0,55. As maiores correlações negativas, em módulo, foram observadas para as espécies Prunus sellowii (ISD1/6), Copaifera langsdorffii (IID2) e Newtonia contorta (IID2), variando entre -0,46 a -0,49.
Estes valores de correlações significativas, em nível de espécie, estão em consonância com valores descritos na literatura. González et al. (2006) ao estudarem a correlação entre o crescimento em diâmetro de Quercus suber, observaram correlação de -0,34 com o IID5. Para Pinus taeda, foram encontradas correlações entre 0,23 e 0,31 entre o crescimento em diâmetro e índices independentes da distância (Daniels et al., 1986). Martins et al. (2011) constataram correlações para crescimento em diâmetro e índices independentes da distância variando entre -0,74 a 0,52 para um híbrido de Eucalyptus.
Cunha (2009), utilizando índices independentes e dependentes da distância, observou correlações entre -0,57 e 0,59 para os melhores índices de competição em relação ao crescimento em área basal da espécie Cedrela odorata, na Floresta Amazônica.
Para o crescimento em altura, Martin e Ek (1984) encontraram valores de correlações inferiores a 0,39 para índices independentes da distância e inferiores a 0,30 para índices dependentes da distância para Pinus resinosa. Martins et al. (2011) encontraram correlações entre índices independentes da distância e crescimento em altura variando entre 0,52 e -0,69 para um híbrido de Eucalyptus. Chassot et al. (2011) observou valores inferiores a -0,64 para correlações entre índices independentes da distância e crescimento em altura em Araucaria angustifolia.
Em relação à mortalidade, Martins et al. (2011) estimaram correlações entre índices independentes da distância e a probabilidade de mortalidade entre -0,28 a 0,62, para um híbrido de Eucalyptus.
De maneira geral, árvores maiores, por estarem no estrato superior das florestas, têm menor competição (Cunha, 2009). Assim, por possuírem melhores condições de iluminação, apresentam maiores taxas de crescimento (King e Maindonald, 1999; Lieberman e Putz, 1987, Costa et al., 2008). Consequentemente, árvores de porte pequeno, localizadas no sub-bosque, têm maior competição e apresentam menores taxas de crescimento (Carvalho, 1992) e maiores probabilidades de mortalidade (Martins et al., 2011). Isso foi observado quando se avaliou o comportamento médio de crescimento em diâmetro e altura e probabilidade de mortalidade para todas as árvores da floresta em conjunto (Figura 3).
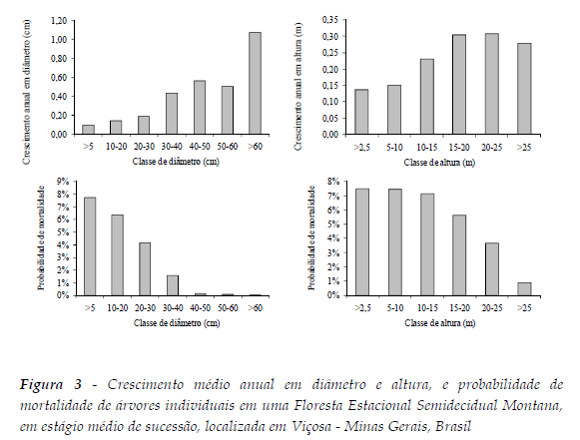
Pode-se observar também, no Quadro 4, que as correlações para as espécies Jacaranda macrantha, Machaerium floridum, Casearia arborea, Mabea fistullifera, Allophylus edulis, Sorocea bonplandii, Endlicheria paniculata, Maytenus robusta apresentaram comportamentos contrários à tendência média de crescimento e/ou mortalidade observada para a média das árvores da floresta (Figura 3), as quais estão destacadas em negrito no quadro. Nestes casos, correlações positivas foram observadas para crescimento (diâmetro ou altura) e negativas para probabilidade de mortalidade para o índice IID5. Para os demais índices, foram verificadas correlações negativas para as variáveis de crescimento e positivas para probabilidade de mortalidade.
Naturalmente, diferentes comportamentos no crescimento e mortalidade podem ser estimulados por outros fatores, como por exemplo, características particulares de grupos ecológicos (Tonini et al., 2008), danos nas copas e presença de cipós (Costa et al., 2008), idade das árvores ou mecanismos de senescência e, até mesmo por fatores desconhecidos (Julian et al., 2001). Pela análise da Figura 4 pode-se constatar o comportamento contrário à tendência média para essas espécies.
Erskine et al. (2005) observaram que algumas espécies de estágios sucessionais iniciais podem apresentar maior crescimento inicial e menor crescimento quanto atingem maior porte. Segundo Tonini et al. (2008) e Costa et al. (2008), essas espécies são adaptadas à colonização e por isso possuem altas taxas de crescimento na fase inicial de desenvolvimento.
Todas as espécies que apresentaram esse comportamento em relação às variáveis de crescimento (dap e Ht) pertencem aos grupos ecológicos das pioneiras ou secundárias iniciais, com exceção da espécie Endlicheria paniculata, que pertence ao grupo das secundárias tardias. Oliveira Júnior (2005), trabalhando em floresta secundária, observou diferença no crescimento da espécie Vochysia maxima, de acordo com o tamanho dos indivíduos, onde indivíduos de menores diâmetros também apresentaram maior crescimento anual em diâmetro do que aqueles de maiores diâmetros, o que normalmente não é esperado.
Não foi observada superioridade de um único índice para as espécies individuais e nem para a floresta como um todo. Isto indica que para cada variável e cada espécie, existe um índice de competição mais eficiente.
No Quadro 5 é apresentado o número de ocorrências de correlações significativas com maiores valores (em módulo) e o valor de superioridade (VS) de cada índice de competição em relação às variáveis de crescimento em dap e Ht, bem como para a Pm.
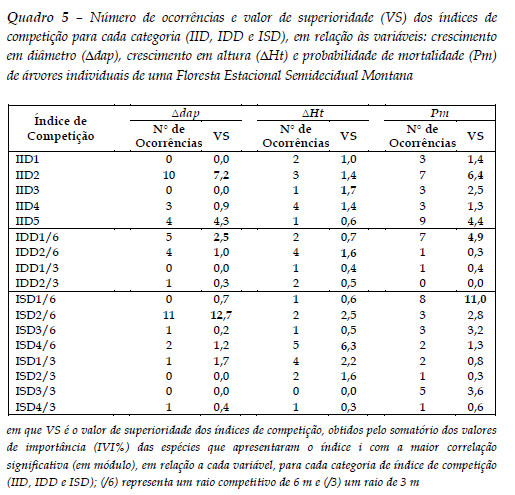
De maneira geral, todas as categorias de índices de competição (IID, IDD e ISD) podem ser utilizadas em modelos de crescimento e mortalidade de árvores individuais. Os índices com maior VS também foram aqueles que apresentaram o maior número de ocorrências dentre as espécies analisadas, exceto para o IID3 para a variável Ht.
Assim sendo, para o crescimento em diâmetro houve superioridade do índice ISD2/6 e para o crescimento em altura, do índice ISD4/6. Para a probabilidade de mortalidade houve superioridade do índice ISD1/6.
Com relação ao raio de competição, para os índices dependentes e semi-independentes, foi observado superioridade para o raio de 6 m para todas as variáveis. Este raio de competição corrobora com estudos que também identificaram o mesmo raio de competição em outras formações florestais (Béland et al., 2003).
Conclusões
Com este estudo pode-se concluir que:
1) Existe correlação significativa entre índices de competição e o crescimento e a mortalidade de árvores no fragmento de Floresta Estacional Semidecidual, em estágio médio de sucessão.
2) Para o local de estudo, os índices semi-independentes da distância são indicados para o estudo de crescimento e de mortalidade regular.
3) O raio de competição de 6 m é indicado para estimação de índices de competição na área de estudo.
Referências
Alig, R.J.; Parks, P.J.; Farrar Jr. R.M.; Vasieviech, J.M., 1984. Regional timber yield and cost information for the South: Modelings techniques. Washington: USDA, 28 pp. [ Links ]
Amaro, M.A., 2010. Quantificação do estoque volumétrico, de biomassa e de carbono em uma floresta estacional semidecidual no município de Viçosa-MG. 2010. 168 f. Tese (Doutorado em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa. [ Links ]
APG III., 2009. An update of APG classification for the orders and families of flowering plants. APG III.Botanical Journal of the Linnean Society (161): 105-121. [ Links ]
Béland, M.; Lussier, J.M.; Bergeron, Y.; Longpré, M.H.; Béland, M., 2003. Structure, spatial distribution and competition in mixed jack pine (Pinus banksiana) stands on clay soils of eastern Canadá. Annals of Forest Science (60): 609-617. [ Links ]
Bigging, G.S.; Dobbertin, M., 1995. Evaluation of competition indices in individual tree growth models. Forest Science 41(2): 360-377. [ Links ]
Bigging, G.S.; Dobbertin, M., 1992. A comparison of distance-dependent competition measures for height and basal area growth of individual conifer tree. Forest Science 38(3): 695-720. [ Links ]
Canham, C.D., LePage, P.T., Coates, K.D., 2004. A neighborhood analysis of canopy tree competition: effects of shading versus crowding. Canadian Journal of Forest Research (34): 778-787. [ Links ]
Carvalho, J.O.P., 1992. Structure and dynamics of logged over Brazilian Amazonian rain forest.215 f. Tese (Doutorado em Ciências Florestais) – University of Oxford, Oxford. [ Links ]
Castro, R.V.O., 2011. Modelagem do crescimento em nível de árvores individuais utilizando redes neurais e autômatos celulares. 80 f. Dissertação (Mestrado em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa. [ Links ]
Chassot, T.; Fleig, F.D.; Finger, C.A.G.; Longhi, S.J., 2011. Modelos de crescimento em diâmetro de árvores individuais de Araucaria angustifolia (Bertol.) Kuntze na Floresta Ombrófila Mista, 2011. Ciência Florestal (21): 303-313. [ Links ]
Coates, K.D., Canham, C.D., LePage, C.T., 2009. Above versus below ground competitive effects and responses of a guild of temperate tree species. Journal of Ecology (97): 118-130. [ Links ]
Contreras, M.A.; Affleck, D., Chung, W., 2011. Evaluating tree competition indices as predictors of basal area increment in western Montana forests. Forest Ecology and Management (262): 1939-1949. [ Links ]
Corrêa, G.F., 1984. Modelo de evolução e mineralogia da fração argila de solos do planalto de Viçosa. 87 f. Dissertação (Mestrado em Solos e Nutrição de Plantas) - Universidade Federal de Viçosa, Viçosa. [ Links ]
Costa, D.H.M.; Silva, J.N.M.; Carvalho, J.O.P., 2008. Crescimento de árvores em uma área de terra firme na floresta nacional do Tapajós após a colheita de madeira. Revista de Ciências Agrárias (50): 63-76. [ Links ]
Cunha, T.A., 2009. Modelagem do incremento de árvores individuais de Cedrela odorata L. na Floresta Amazônica. 88 f. Dissertação (Mestrado em Engenharia Florestal) Universidade Federal de Santa Maria, Santa Maria. [ Links ]
Daniels, R.F.; Burkhart, H.E.; Clason, T.R., 1986. A comparison of competition measures for predicting growth of loblolly pine trees. Canadian Journal of Forest Research (16): 1230-1237. [ Links ]
Della–Flora, J.B., 2001. Modelos de crescimento para árvores singulares de Nectandra megapotamica (Spreng.) Mez. e Ocotea puberula Ness. 60 f. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal de Santa Maria, Santa Maria. [ Links ]
Erskine, P.D.; Lamb, D.; Borschmann, G., 2005. Growth performance and management of a mixed rainforest tree plantation. New Forest (29): 117-234. [ Links ]
Figueiredo, L.T.M., 2011 Dinâmica do estoque de carbono do fuste de árvores de uma Floresta Estacional Semidecidual. 87 f. Dissertação (Mestrado em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa. [ Links ]
Foli, E.G., 1993. Crown dimensions and diameter growth of some tropical mixed forest trees in Ghana. 185 f. Thesis - University of Aberdeen. [ Links ]
Gandolfi, S.; Leitão Filho, H.F.; Bezerra, C.L., 1995. Levantamento florístico e caráter sucessional de espécies arbustivo-arbóreas de uma floresta Mesófila Semidecídua no município de Guarulhos, SP. Revista Brasileira de Biologia 55(4): 753-767. [ Links ]
Glover, G.R.; Hool, J.N., 1979. A basal area ratio predictor of loblolly pine plantation mortality. Forest Science 25(2): 275-282. [ Links ]
González, M.S.; Río, M. del; Cañellas, I.; Montero, G., 2006. Distance independent tree diameter growth model for cork oak stands. Forest Ecology and Management (225): 262-270. [ Links ]
Hamilton Jr., D.A., 1986. A logistic model of mortality in thinned and unthinned mixed conifer stands of Northern Idaho. Forest Science 32(4): 989-1000. [ Links ]
Hasenauer, H.; Monserud, R.A., 1996. A crown ratio model for Austrain forests. Forest Ecology and Management (84):49-60. [ Links ]
Hegyi, F., 1974. A simulation model for managing jack-pine stands. In: FRIES, G. Growth models for tree and stand population. Stockolm: Royal College of forestry, Research note (30): 74-90. [ Links ]
Holmes, M.J.; Reed, D.D., 1991. Competition indices for mixed species northern hardwoods. Forest Science 37(5): 1338-1349. [ Links ]
HYNYNEN, J., 1995. Modelling tree growth for managed stands. The Finnish Forest Research Institute, Research Papers 576, 59 pp. [ Links ]
Inmet – Instituto Nacional de Metrologia, 2012. Banco de Dados Meteorólogicos para Ensino e Pesquisa. Disponível em: http://www.inmet.gov.br/portal/index.php?r=bdmep/bdmep Acesso em: 07/06/2012. [ Links ]
Julian, C.F.; Peter, K.A.; Huiquan, B., 2001. Stochastic structure and individual tree growth models. Forest Ecology and Management (154): 261-276. [ Links ]
King, D.A.; Maindonald, J.H., 1999. Tree architecture in relation to leaf dimensions and tree stature in temperate and tropical rain forests. Journal of Ecology (87): 1012-1024. [ Links ]
Lederman, T., 2010. Evaluating the performance of semi-distance-independent competition indices in predicting the basal area growth of individual trees. Canadian Journal of Forest Research (40): 796-805. [ Links ]
Lieberman, D.; Putz, F.E., 1987. The dynamis of tree populations in tropical forest: a review. Journal of Tropical Ecology (3): 359-366. [ Links ]
Lorimer, C.G., 1983. Test of age-independent competition indices for individual trees in natural hardwood stands. Forest Ecology and Management (6): 343-360. [ Links ]
Martin, G.L.; Ek, A.R., 1984. A comparison of competition measures and growth models for predicting plantation red pine diameter and height growth. Forest Science 30(3): 731-743. [ Links ]
MARTINS, F.B.; SOARES, C.P.B.; LEITE, H.G.; SOUZA, A.L.; CASTRO, R.V.O., 2011. Índices de competição em árvores individuais de eucalipto. Pesquisa Agropecuária Brasileira (46): 1089-1098. [ Links ]
MEIRA NETO, J.A.A.; Martins, f.r., 2000. Estrutura da Mata da Silvicultura, uma Floresta Estacional Semidecidual Montana no município de Viçosa, MG. Revista Árvore 24(2): 151-160. [ Links ]
Monserud, R.A.; Sterba, H., 1996. A basal area increment model for individual trees growing in even-and-uneven-aged forest stands in Austria. Forest Ecology and management (80): 57-80. [ Links ]
Oheimb, G.V.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsche, I.; Yu, M.; Härdtle, W., 2011. Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighbourhood competition. Forest Ecology and Management (261): 499-507.
Oliveira Júnior, J.C.; Dias, H.C.T., 2005. Precipitação efetiva em fragmento secundário da Mata Atlântica. Revista Árvore 29(1): 9-15. [ Links ]
PELTONIEMI, M.; MÄKIPÄÄ, R., 2011. Quantifying distance-independent tree competition for predicting Norway spruce mortality in unmanaged forests. Forest Ecology and Management (261): 30–42. [ Links ]
Pretzsch, H.; Biber, P.; Durský, J., 2002. The single tree-based stand simulator SILVA: construction, application and evaluation. Forest Ecology and Management 162(1): 3-21. [ Links ]
Prodan, M.; Peters, R.; Cox, F.; Real, P., 1997. Mensura forestal. IICA/BMZ/GTZ. 561 pp. [ Links ]
Pukkala, T.; Kolström. T., 1987. Competition indices and the prediction of radial growth in Scots pine. Silva Fennica 21(1): 55-67. [ Links ]
Quicke, H.E., Mehldahl, R.S., Kush, J.S., 1994. Basal area growth of individual trees: a model derived from a regional longleaf pine growth study. Forest Science 40(3): 528-542. [ Links ]
Radtke, P.J., Westfall, J.A., Burkhart, H.E., 2003. Conditioning a distance-dependent competition index to indicate the onset of inter-tree competition. Forest Ecology and Management (175): 17-30. [ Links ]
REZENDE, S.B., 1971. Estudo de cronotoposequência em Viçosa-MG. 71 f. Dissertação (Mestrado em Solos) - Universidade Federal de Viçosa, Viçosa. [ Links ]
Rossi, L.M.B.; Koehler, H.S.; Sanquetta, C.R.; Arce, J.E., 2007. Modelagem de mortalidade em florestas naturais. Revista Floresta 37(2): 275-291. [ Links ]
Sironen, S.; Kangas, A.; Maltamo, A.; Kangas, J., 2002. Estimating individual tree growth with the k-Nearest Neighbour and k-Most Similar Neighbour methods. Silva Fennica 35(4): 453-467. [ Links ]
Stage, A. R., 1973. Prognosis model for stand development. USDA For. Serv. Res. Pap. INT-137. Washington, DC, 32 pp. [ Links ]
Stage, A.R., Ledermann, T., 2008. Effects of competitor spacing in a new class of individual-tree indices of competition: semi-distance-independent indices computed for Bitterlich versus fixed-area plots. Canadian Journal of Forest Research 38(4): 890-898. [ Links ]
Sterba, H.; Blab, A.; Katzensteiner, K., 2002. Adapting an individual tree growth model for Norway Spruce (Picea abies L. Karst.) in pure and mixed species stands. Forest Ecology and Management (159): 101-110. [ Links ]
Tomé, M.; Burkhart, H.E., 1989. Distance-dependent competition measures for predicting growth of individual tree. Forest Science 35(3): 816-831. [ Links ]
Tonini, H.; Oliveira, M.M.C.; Schwengber, D., 2008. Crescimento de espécies nativas da Amazônia submetidas ao plantio no estado de Roraima. Ciência Florestal 18(2): 151-158. [ Links ]
Veloso, H.P.; Rangel Filho, A.L.R.; Lima, J.C.A., 1991. Classificação da vegetação brasileira, adaptada a um sistema universal. Rio de Janeiro: Instituto Brasileiro de Geografia e Estatística (IBGE), 123 p. [ Links ]
Weber, P.; Bugmann, H. Fonti, P.; Rigling, A., 2008. Using a retrospective dynamic competition index to reconstruct forest succession. Forest Ecology and Management 254(1): 96-106. [ Links ]
Wykoff, W.R., 1990. A basal area increment model for individual conifers in the northern rocky mountains. Forest Science 36(4): 1077-1104. [ Links ]
Zhao, D.; Borders, B.; Wilson, M.; Rathbun, S.L., 2006. Modeling neighborhood effects on the growth and survival of individual trees in a natural temperate species rich forest. Ecological Modelling 196(1): 90-102. [ Links ]
Entregue para publicação em novembro de 2013
Aceite para publicação em fevereiro de 2014
Agradecimento
Ao CNPq/Brasil pela bolsa de pesquisa do primeiro autor.
NOTAS
1 Considerando os diferentes raios de vizinhança: 5 índices IID; 4 índices IDD (sendo dois com raio de 6 m e dois com raio de 3 m; e oito índices ISD (sendo quatro com raio de 6 m e dois com raio de 3 m).
