










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkSilva Lusitana
Print version ISSN 0870-6352
Silva Lus. vol.20 no.1-2 Lisboa Dec. 2012
Trapping Monochamus galloprovincialis (Coleoptera: Cerambycidae), Vector of the Pine Wood Nematode, with Pine Allelochemicals, in Portugal
Desenvolvimento de Métodos para a Captura de Monochamus galloprovincialis (Coleoptera: Cerambycidae), Vetor do Nemátode da Madeira do Pinheiro, com Compostos Aleloquímicos do Pinheiro Bravo, em Portugal
Développment de Méthodes pour la Capture de Monochamus galloprovincialis (Coleoptera: Cerambycidae), Vecteur du Nématode du Pin, avec des Composants Alelochimiques du Pin Maritime au Portugal
1 Luis Bonifácio*, 2 Fernando Praias** and 1 Edmundo Sousa***
*Bolseiro de Investigação Doutorado
**Estagiário do curso de Engenharia Florestal
***Investigador Auxiliar
1 INIAV, IP – Instituto Nacional de Investigação Agrária e Veterinária. Quinta do Marquês, Av. da República, 2780-159 OEIRAS
2 Escola Superior Agrária de Castelo Branco. Quinta da Senhora de Mércules. Apartado 119, 6001-909 CASTELO BRANCO
Abstract
Seven different trap models or variations and ten different chemical lures or combinations thereof were compared as to their efficiency to attract and capture the pine sawyer Monochamus galloprovincialis, vector of the Pine Wood Nematode (PWN) (Bursaphelenchus xylophilus) in Portugal, between 2001 and 2004. Traps were made at the INRB laboratories (former EFN), with and without a visual silhouette, with chemical lures (ethanol, α-pinene and/or turpentine) from the maritime pine (Pinus pinaster), the only host of the PWN and its vector in Portugal. They were tested in four successive assays on pine stands; the less efficient were eliminated and the best were accepted for subsequent tests. The best combination was a transparent cross-vane interception trap lured with ethanol and turpentine in separate vials. The implications of the results for control of M. galloprovincialis are discussed.
Key words: Pine wilts disease; maritime pine; pine sawyer; traps; lures
Sumário
Diferentes armadilhas e variações (sete), iscadas com atraentes químicos, foram testadas em Portugal entre 2001 e 2004. O objetivo era verificar a eficiência na atração e captura do longicórnio do pinheiro, Monochamus galloprovincialis. Este é o inseto vetor do nemátodo da madeira do pinheiro (NMP) (Bursaphelenchus xylophilus). Todas as armadilhas testadas foram construídas no laboratório do INRB (ex-Estação Florestal Nacional), sendo algumas de interceção enquanto outras apresentavam silhueta visual. Em todas foram colocados atraentes químicos (etanol, aguarrás e α-pineno) originários do hospedeiro do NMP, o pinheiro-bravo. Foram testadas em quatro ensaios realizados de modo sucessivo, em que as piores combinações foram eliminadas e as melhores admitidas no ensaio seguinte. A melhor combinação era constituída por uma armadilha de painéis cruzados transparentes com um conjunto atrativo constituído por etanol e aguarrás, apresentados em recipientes separados. Finalmente são discutidas as implicações dos resultados obtidos no controlo das populações de M. galloprovincialis.
Palavras-chave: Bursaphelenchus xylophilus; armadilhas; atraentes químicos; pinheiro-bravo
Résumé
Différentes pièges et variations (sept), avec des attractives chimiques, ont été testés entre 2001 et 2004 pour leur efficacité à attirer et capturer Monochamus galloprovincialis, insecte vecteur du nématode du pin (PWN) (Bursaphelenchus xylophilus) au Portugal. Toutes les pièges ont était fabriquées dans le laboratoire de l'INRB(anciennement Station Forestière Nationale), quelques-unes d'interception tandis que d'autres avaient silhouette pour contact visuel de l'insecte. Toutes avaient des produits chimiques attractifs (éthanol, essence de térébenthine et α-pinène) provenant de l'hôte du nématode de pin, le pin maritime, et ont été testées dans quatre essais réalisés en succession, dans laquelle la pire des combinaisons a été éliminée et la meilleure admis à l'essai suivante. La meilleure combinaison finale était composée d'un piège à panneaux transparents plié avec des attractantes constitué d'éthanol et essence de térébenthine dans des conteneurs individuels.
Mots clés: Maladie du nématode du pin; longicorne du pin; pin maritime; piège; attractante chimique
Introduction
The pine wood nematode (PWN) Bursaphelenchus xylophilus (Steiner & Bührer) Nickle, the causal agent of pine wilt disease (MOTA et al., 1999), was discovered in Portugal in 1999. Soon after, the main study allowed identification of its unique vector, the native secondary forest insect Monochamus galloprovincialis (Olivier) (SOUSA et al., 2001).
Since the detection of the first infected pines, enormous efforts have been made to control the disease, namely by cutting and destroying the infested trees before the exit of the insect vector and by capturing the insect vector during its flight period.
For this last purpose, early assays used traps, made at the INRB Laboratory based on descriptions from publications, with kairomone lures from the maritime pine tree host (ethanol, turpentine and α-pinene), as tested in other countries where PWN causes mortality: Asia (Japan) and exotic pines in the USA.
Many traps were developed to catch conifer bark and wood-boring beetles. They can be divided into four different types: transparent flight interception (i.e. cross-vane traps) (BILLINGS and CAMERON, 1984), silhouette or barrier traps (i.e. multi-funnel traps) (LINDGREN, 1983; FATZINGER, 1985), sticky traps (IKEDA et al., 1980) and vertical stove-pipe traps (CHÉNIER and PHILOGÈNE, 1989b).
Different traps and lures to catch the PWN Monochamus vector have already been studied, mainly in USA and Japan. As is the case with most cerambicids, communication in the Monochamus genus is based on chemical stimuli, flying in the host volatiles' plume (TUNSET et al., 1988; ALLISON et al., 2004). These are namely terpenes (CHÉNIER and PHILOGÈNE, 1989a; DYER and SEABROOK, 1978; ZHANG et al., 1993) and ethanol (CHÉNIER and PHILOGÈNE, 1989b). Sometimes they also need visual clues (SEYBERT and GARA, 1970; VASECHKO, 1978; NIEMEYER, 1985) and mechanical search after landing (DICKE and GROSTAL, 2001).
Cerambicids are sensitive to volatiles produced by the host, using the chemical clues for feeding, mating and egg-laying. Host selection might initially be based on chemical odors and its acceptance regulated by tasting, all depending on the volatile terpene content of the wood (HANKS, 1999; FACCOLI et al., 2005; GINZEL and HANKS, 2003).
The objectives of this paper are to study the response of M. galloprovincialis to the host semiochemical odors, to turpentine and ethanol and to test several trap designs (LINDGREN, 1983; FATZINGER, 1985).
Materials and methods
Traps and lures developed to catch M. galloprovincialis were tested in the initially affected area of Setúbal Península, at Tróia (2001 and 2002) and Comporta (2004). It was necessary to displace the assays further south from Comporta because the insect population levels at Tróia decreased due to the success of eradication procedures, with an impact on trap captures.
All assays occurred from July to September, in order to overlap with the flight period of M. galloprovincialis adults (NAVES et al., 2008). Traps were visited weekly to collect captured bark beetles and wood borers and lures were renewed when vials were found to be almost empty.
Traps and/or lures with the best results in one assay were used again in the following assay against new sets. Table 1 presents the list of materials used in the different assays. All traps used in the work described in this paper were built at the INRB Laboratory. In the field, traps were hung high in the lower canopy of adult pines. This procedure was followed in order to improve trapping efficiency by placing them at the height of M. galloprovincialis flight and trunk colonization.
Table 1 – Traps and lures used in assays for the capture of Monochamus galloprovincialis
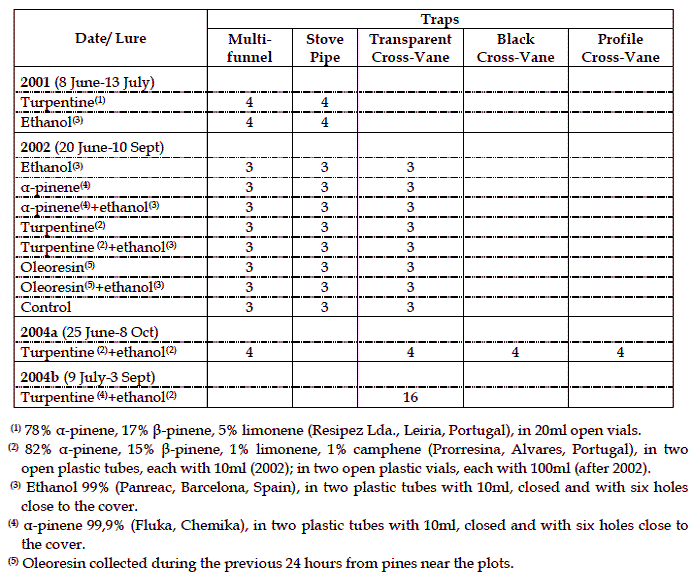
The multi-funnel trap was based on Lindgren's trap (1983) using nine green plastic funnels (16cm high and 20cm wide) with a narrow end cut. The bottom funnel was glued to a 50cl plastic cup containing a DDVP insecticide strip (Figure 1a).

Figure 1 – Traps used in assays to capture Monochamus galloprovincialis at Tróia and Comporta. a- multi-funnel; b- stove pipe; c to e – cross vane traps
The stove pipe trap was based on the Fatzinger trap (1985), using a black PVC pipe 80cm high and 15cm wide, attached on the bottom to a black, round 50cm-wide trash bin cover. The whole setup was covered with the entomologic glue Temocid (Quelmer, Lisboa, Portugal). Attractants were hung inside the tube and to allow dispersal of odors, two rings of holes were drilled into the tube at 25cm from each end. To prevent the capture of small birds or bats in the glue a 5cm-wide metallic net was attached to the border of the trash bin cover (Figure 1b).
The transparent cross vane trap was based on traps described by WILKENING et al. (1981) and a Japanese commercial trap for M. alternatus (NAKAMURA et al., 1999). The trap consisted of two acrylic plates, 30cm wide and 40cm high, cross-connected in the center through 15cm slits. A 30cm funnel was hung under the vanes to lead the insect that crashed into them to a 50cl plastic vial glued at the funnel end and containing a DDVP insecticide strip (Figure 1c).
The first variation of the cross vane trap was completely black, as described by McIntosh et al. (2001) (Figure 1d), and the second had a 10cm vertical black band at the middle, where the vanes cross (GROOT and NOTT, 2001) (Figure 1e).
Experimental design
The assay done in 2002, in Tróia pine stands, compared three different traps (multi-funnel, stove pipe and transparent cross vane), seven allelochemical combinations (oleoresin, turpentine, α-pinene) and a control (no lure).
Eight groups were established with 100m between them, with one trap of each type having the same attractant, placed 50m apart. Every week attractants were replaced by new ones of a different kind following a clockwise order (Figure 2).
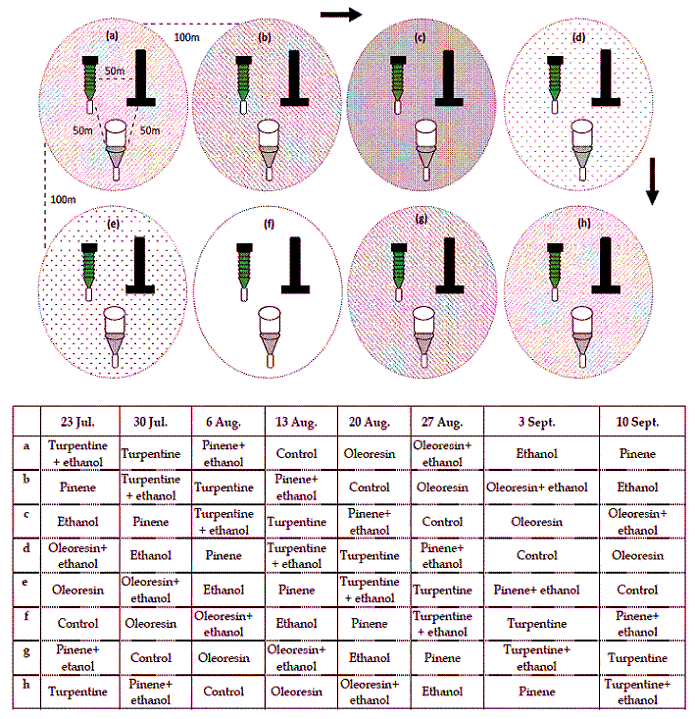
Figure 2 – Experimental design used to test traps and chemical lures to capture Monochamus galloprovincialis, with 3 simultaneous replications on the field. Arrows indicate the direction of clockwise rotation of the lures between groups of traps, according to the time sequence described in the table
This way, after eight weeks all lures had been used in all traps and the assay ended. This design was repeated three times and as a result this assay used 72 traps simultaneously.
In all the other experiments the design used was the Latin square, with traps placed 100m from each other and, in each line or column of traps, all tested variables were represented without repetition.
In 2001 multi-funnel and stove pipe traps were tested, lured with turpentine+ethanol and pinene+ethanol. In a first assay in 2004 the three variations of the cross-vane and multi-funnel traps were compared, all lured with turpentine+ethanol. Finally, in a second 2004 assay, 16 transparent cross vane traps were used to test four combinations of turpentine and ethanol, mixed at different ratios (1:1, 1:2 and 2:1) as well as in separate vials.
Statistical analysis
In all the assays' results one-way ANOVA was applied to assess the effect of each variable: chemical lure, trap and time. Afterward, variables were tested together using multiple ANOVA, where interaction between fixed (trap and lure) and random (time) factors was evaluated for its effectiveness in attracting and capturing M. galloprovincialis adults.
Since frequently less than 10 beetles were captured and some traps did not even capture a single insect, results were transformed by ![]() , also used to assure data normality and homocedasticity (ROELOFS and CARDE, 1977; ZAR, 1984).
, also used to assure data normality and homocedasticity (ROELOFS and CARDE, 1977; ZAR, 1984).
When statistically significant differences were obtained a post-hoc Fisher Least Square Difference was adjusted, because it is considered to be more robust than other available tests such as Tukey HSD, Newman-Keuls, Bonferroni, Scheffé and Duncan (ZAR, 1984).
Results
The time pattern of M. galloprovincialis captures was studied for the 2002 assay and a concentration was observed in the first two weeks, with 18 and 17 beetles respectively (79,5% of 44 insects caught during the whole test). Variance analysis revealed significant concentration during these weeks and a weak contribution during the third and eighth weeks (Figure 3). The reason for the sudden decrease during the third week was due to unstable weather observed (rain and strong winds) that surely conditioned the beetles' activity. Disregarding this week the pattern showed a slow decrease towards the end of the assay, in late September, when the emergence period of M. galloprovincialis ends.
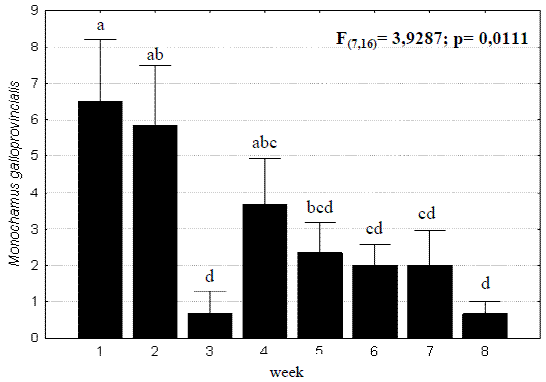
Figure 3 – Weekly pattern of total Monochamus galloprovincialis captures observed in the 2002 assay. Lines are standard errors associated to means (in black bars) and letters are Homogeneous LSD groups
Traps
The first assay to study the effect of trap design was conducted in 2001, when 38 adult M. galloprovincialis were captured by two kinds of traps, mainly the stove-pipe trap (27 specimens against 11 in the multi-funnel trap) but no significant statistical difference was found (F(6,1)= 0,3139; p= 0,5956) (Figure 4).
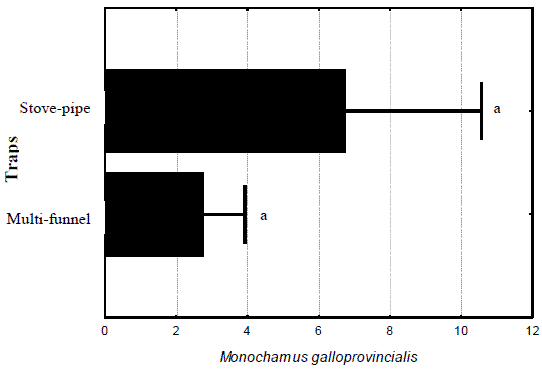
Figure 4 – Total captures of Monochamus galloprovincialis adults obtained per trap in two different traps tested in 2001. Lines are standard errors associated to means (in black bars) and letters are Homogeneous LSD groups
In the 2002 assay, concerning the performance of different traps, the multi-funnel traps did not catch any M. galloprovincialis, therefore the ANOVA analysis showed significant differences in relation to the other two trap types, that showed similar capabilities (F(2,69)= 11,2129, p< 0,0001) (Figure 5).
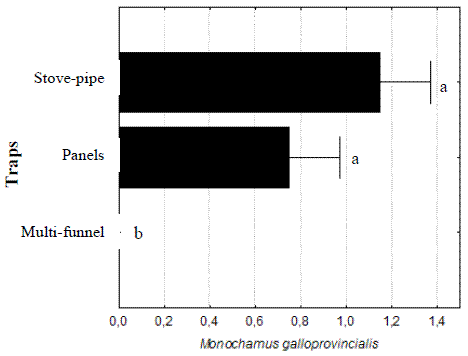
Figure 5 – Total numbers of Monochamus galloprovincialis caught per trap and per week in assay of 2002. Lines are standard errors associated to means (in black bars) and letters are Homogeneous LSD groups
In the trap assay done in 2004 the introduction of a visual effect in cross-vane traps was tested. Once again the multi-funnel trap gave the worst results (though having captured four beetles during the 16 weeks of the test period).
Transparent cross-vane traps captured 24 M. galloprovincialis beetles, followed by the completely black traps with 23 beetles; profile traps caught 26 beetles (although almost half of them were caught during the first week in a single trap). Transparent cross-vane traps showed more constant captures distributed between the four traps used and along the weeks of the assay.
One-way ANOVA analysis reported statistically significant differences in total captures (F(3,12)= 2,7594; p= 0,0882) and of males (F(3,12)= 3,6371; p= 0,0449), with negative displacement of multi-funnel in relation to cross-vane traps (Figure 6). Females were captured in lesser numbers (n = 31) and equally in all kinds of traps (F(3,12)= 1,4412; p= 0,2796).
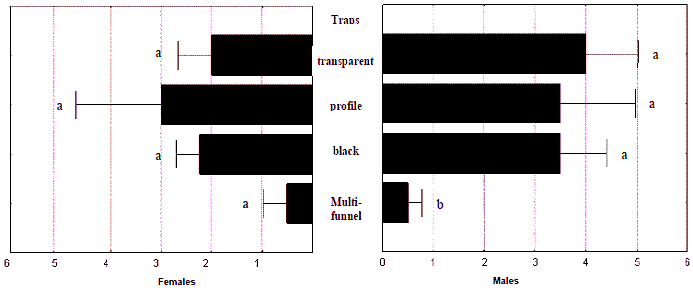
Figure 6 – Total numbers of Monochamus galloprovincialis caught per trap and per week in assay of 2004a. Lines are standard errors associated to means (in black bars) and letters are Homogeneous LSD groups
Lures
During the 2001 assay, M. galloprovincialis were only caught in traps lured with ethanol and pinene; none were caught in traps with turpentine.
The most consistent result obtained in the 2002 assay was the synergistic effect of ethanol when combined with other compounds; the M. galloprovincialis captures obtained were multiplied by 1,7 with α-pinene, by 2,3 with oleoresin and by 7,7 when combined with turpentine, giving a strong contribution to the statistical differences observed with ANOVA analysis (F(7,64)= 2,9065, p= 0,0106). Ethanol alone proved to be more attractive than turpentine or oleoresin placed in isolation (Figure 7).
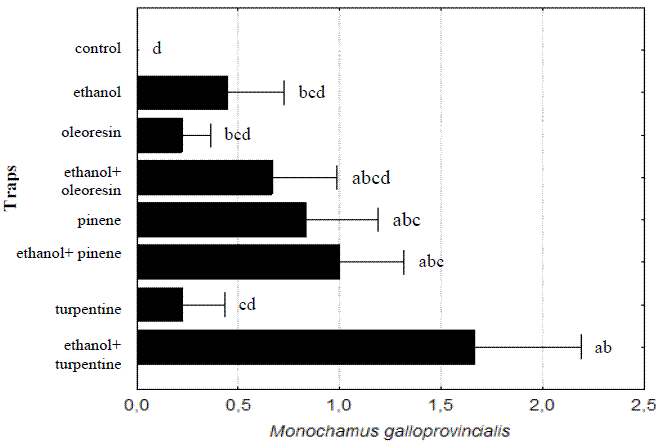
Figure 7 - Total numbers of Monochamus galloprovincialis caught per trap and per week in the 2002 assay. Lines are standard errors associated to means (in black bars) and letters are Homogeneous LSD groups
The combined effects of traps and lures decreased previous observed differences (F(14,48)= 1,3116; p= 0,2359). However, the best results of all traps were always obtained when used with ethanol and turpentine (a total of eight beetles in transparent panels and seven beetles in stove-pipe).
In the other assay done in 2004 with the purpose of clarifying the best way to present ethanol and turpentine, when these attractants were placed separately, more than half of the total M. galloprovincialis beetles were caught (54 of 103 beetles).
Statistical differences were only found with trapped males (F(3,12)= 3,3813; p= 0,0543) because there was a concentration of females in one trap with separated chemicals (17 out of 25 total females caught), resulting in a big standard error, either in relation to female trapping (F(3,12)= 1,3619; p= 0,3013) or total trapping (F(3,12)= 2,5865; p= 0,1016).
Different mixtures of ethanol and turpentine gave similar results (Figure 8).
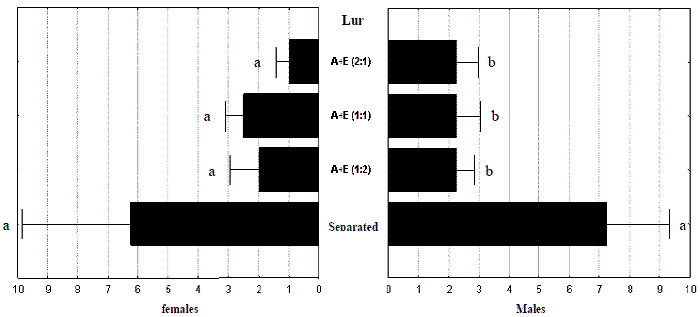
Figure 8 – Total numbers of Monochamus galloprovincialis caught per trap and per week in the 2004b assay. Turpentine (A) and Ethanol (E) were placed separately and in mixtures at different proportions (2:1; 1:2 and 1:1). Lines are standard errors associated to means (in black bars) and letters are Homogeneous LSD groups
Considering other wood-boring species, in the 2002 assay a total of 240 cerambicids, 47 buprestids and 20 curculionids were caught. Besides the 44 M. galloprovincialis specimens, the other cerambicids caught were 95 Ahropalus syriacus, 7 A. ferus, 32 S. buprestoides, 50 P. perroudi, 11 A. griseus and 1 Trichoferus sp..
In our assays most of the M. galloprovincialis beetles were captured in transparent cross-vane traps, so a visual stimulus is not important for catching M. galloprovincialis; odors therefore play the major role, as referred by PAJARES et al. (2004).
Silhouette and stove-pipe traps have a strong visual effect which is important to many species that need visual stimuli to stop their flight and land. Flight interception traps only rely on the attractive ability of the lure and have proven to be efficient in catching large forest insects (YOUNAN and HAIN, 1982). Most North American Monochamus species such as M. titillator, M. carolinensis (FATZINGER, 1985) and M. notatus (CHENIÉR and PHILOGÈNE, 1989b) were caught in higher numbers in traps with a black vertical silhouette than in interception or multi-funnel traps, although in some situations no statistical differences were found (CHÉNIER and PHILOGÈNE, 1989a; GROOT and NOTT, 2001; 2003; MOREWOODet al., 2002). On the other hand, GROOT and NOTT (2001; 2003) and McINTOSH et al. (2001) found differences between Monochamus species, where visual stimuli is important for M. scutellatus but not for M. mutator.
In our assays, multi-funnel traps always showed the worst results when compared with the others. Multi-funnel traps are useful for monitoring but not efficient for pine sawyer population limitation (NAKAMURA, 2003; LOYD et al., 2004), as the Japanese trap for M. alternatus (SHIBATA et al., 1996; SAKAKIBARA et al., 1998; NAKAMURA et al., 1999; NAKAMURA and SONÉ, 2004). The use of sticky traps, such as the stove-pipe, although they had proven to be efficient in catching M. galloprovincialis, is not practical for large-scale trapping and it is impossible to extract nematodes, needed for infestation assessment within the affected area.
Since in many of the assays no statistical differences were found between the traps used, the attractive effect of chemical compounds is apparently the most important variable for catching M. galloprovincialis.
The first two assays used different turpentines and the 2001 turpentine was produced from a mixture of oleoresin from P. pinaster andP. pinea with higher limonene content. This difference must have had a strong repellent effect on M. galloprovincialis. Several studies report that limonene has an opposite effect to the attraction produced by α-pinene on H. abietis (LINDGREN et al., 1996; KLEPZIG and SCHLYTER, 1999), inhibits egg-laying of pine processionay moth (TIBERI et al., 1999; ZHANG et al., 2003), is toxic to many bark beetles (BYERS, 1995) and is a feeding deterrent for the larvae of M. alternatus (FAN and SUN, 2006).
The strong synergistic effect of ethanol when combined with oleoresin compounds found in our results was also reported by other authors and may be related to the fact that ethanol is a subproduct of microbial activity inside tree hosts and is interpreted by secondary insects as a sign of a weakened host (IKEDA and ODA, 1980; BYERS, 1989a; b; SATO and MAETO, 2006).
The most abundant terpene of the pines' oleoresin is α-pinene (around 80% in Portuguese P. pinaster) (PESTANA, 1993); when combined with ethanol, it has proven to be very attractive to bark and wood-boring beetles (BORDEN et al., 1982; SCHROEDER and LINDELOW, 1989; BYERS, 1992).
This time pattern observed in our assays, with a strong effect in the first two weeks, was also reported for M. alternatus adults and related to high initial sensitivity to α-pinene, becoming less effective as the insect matures sexually (SAKAI et al., 1992).
However, we obtained better results with turpentine than with α-pinene alone, as in other studies with wood-boring beetles (FATZINGER, 1985; IKEDA et al., 1986; PHILLIPS et al., 1988; CHÉNIER and PHILOGÈNE, 1989b; ALLISON et al., 2001) and pine processionary moth (ZHANG et al., 2003). This reveals that M. galloprovincialis may be sensitive to a minor compound of oleoresin, still to be isolated.
The trapping of M. galloprovincialis with lures based on allelochemicals produced by the pine tree host, inside the area affected by the pine wood nematode, has a limited effect because all year round many diseased trees can be found highly attractive to beetles.
To improve the efficiency of traps in capturing M. galloprovincialis in order to serve as an important tool for pine sawyer population regulation and therefore reduce contamination by the pine wood nematode B. xylophilus, it is essential to find non-tree host odors that would easily be detected by the beetles, even in a high-mortality pine stand.
References
ALLISON, J.D., BORDEN, J.H., McINTOSH, R.L., GROOT, P., GRIES, R., 2001. Kairomonal responses by four Monochamus species (Coleoptera: Cerambycidae) to bark beetle pheromones. J. Chem. Ecol. 27: 633-646. [ Links ]
ALLISON, J.D., BORDEN, J.H., SEYBOLD, S.J., 2004. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 14: 123–150. [ Links ]
BILLINGS, R.F., CAMERON, R.S., 1984. Kairomonal responses of Coleoptera, Monochamus titilator (Cerambycidae), Thanasimus dubius (Cleridae), and Temnochila virescens (Trogositidae) to behavioural chemicals of southern pine bark beetles (Coleoptera: Scolytidae). Environ. Entomol. 13: 1542-1548. [ Links ]
BORDEN, J.H., KING, C.J., LINDGREN, S., CHONG, L., GRAY, D.R., OEHLSCHLAGER, A.C., SLESSOR, K.L., PIERCE JR., H.D., 1982. Variation in response of Trypodendron lineatum from two continents to semiochemicals and trap form. Environ. Entomol. 11: 403-408. [ Links ]
BYERS, J.A., 1989a. Chemical ecology of bark beetles. Experientia 45: 271-283. [ Links ]
BYERS, J.A., 1989b. Behavioural mechanisms involved in reducing competition in bark beetles. Holartic Ecology 12: 466-476. [ Links ]
BYERS, J.A., 1992. Attraction of bark beetles, Tomicus piniperda, Hylurgops palliatus and Trypodendron domesticum and other insects to short-chain alcohols and monoterpenes. J. Chem. Ecol. 18(12): 2385-2402. [ Links ]
BYERS, J.A., 1995. Host-tree chemistry affecting colonization in bark beetles. in Cardé & Bell Eds. Chemical ecology of insects 2: 154-213. [ Links ]
CHÉNIER, J.V.R., PHILOGÈNE, B.J.R., 1989a. Field responses of certain forest Coleoptera to conifer monoterpenes and ethanol. J. Chem. Ecology 15(6):1729-1745. [ Links ]
CHÉNIER, J.V.R., PHILOGÈNE, B.J.R., 1989b. Evaluation of three trap designs for the capture of conifer-feeding beetles and other forest Coleoptera. Can. Ent. 121: 159-167. [ Links ]
DICKE, M., GROSTAL, P., 2001. Chemical detection of natural enemies by arthropods: an ecological perspective. Ann. Rev. Ecol. Syst. 32: 1-23. [ Links ]
DYER, L.J., SEABROOK, W.D., 1978. Some aspects of oviposition site selection in Monochamus notatus e M. scutellatus (Coleoptera: Cerambycidae). J. Chem. Ecology 4(2): 199–210. [ Links ]
FACCOLI, M., BLAZENEC, M., SCHLYTER, F., 2005. Feeding responses to host and nonhost compounds by males and females of the spruce bark beetle Ips typographus in a tunnelling microassay. J. Chem. Ecol. 31(4): 745-759. [ Links ]
FAN, J.T., SUN, J.H., 2006. Influences of host volatiles on feeding behaviour of the Japanese pine sawyer, Monochamus alternatus. J. Appl. Entomol. 130(4): 238-244. [ Links ]
FATZINGER, C.W., 1985. Attraction of black turpentine beetle (Coleoptera: Scolytidae) and other forest Coleoptera to turpentine-baited traps. Environ. Entomol. 14(6): 768-775. [ Links ]
GINZEL, M.D., HANKS, L.M., 2003. Contact pheromones as mate recognition cues of four species of Longhorned beetles (Coleoptera: Cerambycidae). J. Insect Behaviour 16(2): 181-186. [ Links ]
GROOT, P., NOTT, R., 2001. Evaluation of traps of six different designs to capture pine sawyer beetles (Coleoptera: Cerambycidae). Agric. For. Entomol. 3: 107-111. [ Links ]
GROOT, P., NOTT, R., 2003. Response of Monochamus (Col., Cerambycidae) and some Buprestidae to flight intercept traps. J. Appl. Entomol. 127: 548-552. [ Links ]
HANKS, L.M., 1999. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 44: 483-505. [ Links ]
IKEDA, T., ODA, K., 1980. The occurrence of attractiveness for Monochamus alternatus Hope (Coleoptera: Cerambycidae) in nematode-infected pine trees. J. Jap. For. Soc. 62(11): 432-434. [ Links ]
IKEDA, T., ODA, K., YAMANE, A., ENDA, N., 1980. Volatiles from pine logs as the attractants for the Japanese Pine Sawyer Monochamus alternatus Hope (Coleoptera: Cerambycidae). J. Jap. For. Soc. 62(4): 150-152. [ Links ]
IKEDA, T., YAMANE, A., ENDA, N., ODA, K., MAKIHARA, H., ITO, K., ÔKOCHI, I., 1986. Attractiveness of each volatile components of felled pine trees for Monochamus alternatus (Coleoptera: Cerambycidae). J. Jap. For. Soc. 68: 15-19. [ Links ]
KLEPZIG, K.D., SCHLYTER, F., 1999. Laboratory evaluation of plant-derived antifeedants against the pine weevil Hylobius abietis (Coleoptera: Curculionidae). J. Econ. Entomol. 92(3): 644-650. [ Links ]
LINDGREN, B.S., 1983. A multiple funnel trap for Scolitid beetles (Coleoptera). Can. Ent. 115: 299-302. [ Links ]
LINDGREN, B.S., NORDLANDER, G., BIRGERSSON, G., 1996. Feeding deterrence of verbenone to the pine weevil, Hylobius abietis (L.) (Col., Curculionidae). J. Appl. Entomol. 120: 397–403. [ Links ]
LLODY, F., NAKAMURA, K., LINIT, M., 2004. Seasonality of forest pest and beneficial insects attracted to alpha–pinene / ethanol baited Lindgren traps in a study to monitor Monochamus carolinensis in Missouri pine stands. in Proc. Entomol. Soc. America Ann. Meeting: 1 pp. [ Links ].
McINTOSH, R.L., KATINIC, P.J., ALLISON, J.D., BORDEN, J.H., DOWNEY, D.L., 2001. Comparative efficacy of five types of trap for woodborers in the Cerambycidae, Buprestidae and Siricidae. Agricultural and Forest Entomology 3: 113-120. [ Links ]
MOREWOOD, W.D., HELN, K.E., KATANIC, P.J., BORDEN, J.H., 2002. An improved trap for large wood-boring insects with special reference to Monochamus scutellatus (Coleoptera: Cerambycidae). Can. J. For. Res. 32: 519-525. [ Links ]
MOTA, M.M., BRAASCH, H., BRAVO, M.A., PENAS, A.C., BURGERMEISTER, W., METGE, K., SOUSA, E., 1999. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1(7-8): 727-734. [ Links ]
NAKAMURA, K., 2003. Monitoring of the Japanese pine sawyer, Monochamus alternatus and its load of the pinewood nematode, Bursaphelenchus xylophilus, with a commercial attraction trap modified for live capture. in Korea-Japan Joint Conf. Appl. Entomol. Zool.: 180. [ Links ]
NAKAMURA, K., SONE, K., 2004. Modification of capturing bucket of the attraction trap for live trapping of Monochamus alternatus adults to prevent escaping beetles. Kyushu J. Forest. Res. 57: 110-112. [ Links ]
NAKAMURA, K., SONÉ, K., OOKUMA, H., 1999. Modification of a commercial attraction trap for live trapping of Monochamus alternatus (Hope) adults (Coleoptera; Cerambycidae). Appl. Entomol. Zool. 43: 55-59. [ Links ]
NAVES, P.M., SOUSA, E., RODRIGUES, J.M., 2008. Biology of Monochamus galloprovincialis (Coleoptera: Cerambycidae) in the pine wilt disease affected zone, Southern Portugal. Silva Lusitana 16(2): 133-148. [ Links ]
NIEMEYER, H., 1985. Field response of Ips typographus (Col., Scolytidae) to different trap structures and white versus black flight barriers. Z. Ang. Ent. 99: 44-51. [ Links ]
PAJARES, J.A., IBEAS, F., DIEZ, J.J., GALLEGO, D., 2004. Attractive responses by Monochamus galloprovinciallis (Col., Cerambycidae) to host and bark beetle semiochemicals. J. Appl. Entomol. 128: 633-638. [ Links ]
PESTANA, M., 1993. Caracterização da aguarrás proveniente de cinco espécies de pinheiro. Silva Lusitana 1(1): 109-112. [ Links ]
PHILLIPS, T.W., WILKENING, A.J., ATKINSON, T.H., NATION, J.L., WILKINSON, R.C., FOLTZ, J.L., 1988. Synergism of turpentine and ethanol as attractants for certain pine-feeding beetles (Coleoptera). Environ. Entomol. 17(3): 456-462. [ Links ]
ROELOFS, W.L., CARDE, R.T., 1977. Responses of Lepidoptera to synthetic sex phero-mone chemicals and their analogues. Ann. Rev. Entomol. 22: 377-405. [ Links ]
SAKAI, M., KOYAMA, T., OHARA, T., HONDA, Y., YAMAMOTO, H., YAMASAKI, T., 1992. Host plant terpenes affecting age-related differences in olfactory orientation of the cerambycid beetle Monochamus alternatus Hope. Tech. Bull. Fac. Agr. Kagawa Univ. 44: 107–112. [ Links ]
SAKAKIBARA, Y., KIKUMA, A., IWATA, R., YAMANE, A., 1998. Performances of four chemicals with floral scents as attractants for longicorn beetles (Coleoptera: Cerambycidae) in a broadleaved forest. J. For. Res. 3: 221–224. [ Links ]
SATO, S., MAETO, K., 2006. Attraction of female Japanese horntail Urocerus japonicus (Hymenoptera: Siricidae) to α-pinene. Appl. Entomol. Zool. 41(2): 317-323. [ Links ]
SCHROEDER, L.M., LINDELOW, A., 1989. Attraction of scolitids and associated beetles by different absolute amounts and proportions of α-pinene and ethanol. J. Chem. Ecology. 15(3): 807-817. [ Links ]
SEYBERT, J.P., GARA, R.I., 1970. Notes on flight and host-selection behaviour of the pine engraver, Ips pini (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 63: 947-950. [ Links ]
SHIBATA, E., SATO, S., SAKURATANI, Y., SUGIMOTO, T., KIMURA, F., ITO, F., 1996. Cerambycid beetles (Coleoptera) lured to chemicals in the forests of Nara Prefecture, central Japan. Ann. Entomol. Soc. Am.89: 835-842. [ Links ]
SOUSA, E., BRAVO, M.A., PIRES, J., NAVES, P., PENAS, A.C., BONIFÁCIO, L., MOTA, M., 2001. Bursaphelenchus xyliphilus (Nematoda; Aphelenchoididae) associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology. 3(1): 89-91. [ Links ]
TIBERI, R., NICCOLI, A., CURINI, M., EPIFANO, F., MARCOTULLIO, M.C., ROSATI, O., 1999. The role of the monoterpene composition in Pinus spp. Needles, in host selection by the pine processionary caterpillar, Thaumetopoea pityocampa. Phytoparasitica 27(4): 263-272. [ Links ]
TUNSET, K., NILSSEN, A.C., ANDERSEN, J., 1988. A new trap design for primary attraction of bark beetles and bark weevils (Col., Scolytidae and Curculionidae). J. Appl. Entomol. 106: 226-269. [ Links ]
VASECHKO, G.I., 1978. Host selection by some bark beetles (Coleoptera: Scolytidae). I - Studies of primary attraction with chemical stimuli. J. Appl. Ent. 85: 66-76. [ Links ]
WILKENING, A.J., FOLTZ, J.L., ATKINSON, T.H., CONNOR, M.D., 1981. An omnidirectional flight trap for ascending and descending insects. Can. Ent. 113: 453-455. [ Links ]
YOUNAN, E.G., HAIN, F.P., 1982. Evaluation of five trap designs for sampling insects associated with severed pines. Can. Ent. 114: 789-796. [ Links ]
ZAR, J.H., 1984. Biostatistical analysis. 2nd Edition. Prentice-Hall Int. Inc., New Jersey. [ Links ]
ZHANG, Q.H., BYERS, J., ZHANG, X.D., 1993. Influence of bark thickness, trunk diameter and height on reproduction of the Longhorned beetle Monochamus sutor (Col., Cerambycidae) in burned larch and pine. J. Appl. Entomol. 115: 145-154. [ Links ]
ZHANG, Q.H., SCHLYTER, F., BATTISTI, A., BIRGERSSON, G., ANDERSON, P., 2003. Electrophysiological responses of Thaumetopoea pityocampa females to host volatiles: implications for host selection of active and inactive terpenes. J. Pest Science 76: 103-107. [ Links ]
Acknowledgements
The work presented was partially funded by the European Union Fifth Framework, project QLK5-CT-2002-00672-PHRAME (Development of improved pest risk analysis techniques for quarantine pests, using pinewood nematode, B. xylophilus, in Portugal as a model system). We thank Eng.ª Célia Ferreira of TróiaResort and Eng. José Manuel Rodrigues of ICNF (Instituto da Conservação da Natureza e Florestas) for their assistance in establishing the traps at Tróia and Comporta, respectively.
The author would like to acknowledge Dr. Pedro Naves for critical review of the final draft.
(MEDINSECT, 2008)
