










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkSilva Lusitana
versão impressa ISSN 0870-6352
Silva Lus. vol.20 no.1-2 Lisboa dez. 2012
Evaluation of the Potential of Sequestration of Atmospheric Carbon in a Portuguese Eucalypt Stand after Felling
Evolução do Potencial de Sequestro do Carbono Atmosférico num Eucaliptal após o Corte
Évaluation du Potentiel de Séquestre du Carbone Atmosphérique dans un Peuplement d'Eucalyptus après la Coupe
Abel Rodrigues* and Gabriel Pita**
*Senior Researcher
Instituto Nacional de Investigação Agrária e Veterinária. UEISSAFSV. Av. da República. Quinta do Marquês, 2780-159 OEIRAS
**Assistant Professor
Instituto Superior Técnico. Av. Rovisco Pais, 1049-001 LISBOA
Abstract
In Portugal, eucalypt (Eucalyptus globulus Labill.) stands aimed mainly for pulp production, occupy an area of 739,515ha, corresponding to 23% of total forest area. The main objective of this work was to report the recovery of seasonal pattern of GPP in a eucalypt site in Pegões (Southern Portugal) after a felling carried out in October - November 2006. The site was part of a 300 ha eucalypt stand, located in Herdade da Espirra, with a density of about 1100 trees/ha. The stand was intensively managed as a coppice under a twelve year productive cycle and characterized by a 12-month growing period. Carbon fluxes data in this site have been measured by eddy covariance in the period 2002-2010. A prolonged two stage drought in 2004 and 2005, when rainfall was reduced to values of 50% below the long-term mean term, and the clear cutting, followed by the start of a new production cycle, changed the carbon sink ability of eucalypt stand. In the period prior to cutting GPP (2204 g m-2) was maximum in 2002 decreasing to a minimum of 1255.1 g m-2 in 2005 at the peak of the drought. Eight months after the felling young shoots emerging from stumps grew strongly and the new trees reached an average 8m height in 2010. In June 2007 the eucalypt stand recovered its carbon sink capacity with an annual GPP of 1621.6 g m-2 in 2010. Seasonal patterns of GPP in 2008 and 2009 with maximum carbon uptake occurring from mid-February to mid-October were almost opposite to that of the period before the felling when leaf stomatal control prevented major water losses under summer water stresses. The fast recovery of the young eucalypt stand was due to the maintenance of a viable and mature root system. In 2010 GPP monthly variation showed a pattern analogous to the period prior to drought.
Key words: Carbon uptake; stomatal control; seasonality; coppice; mature root system
Sumário
Em Portugal os povoamentos de eucalipto (Eucalyptus globulus Labill.) destinados principalmente à produção de pasta de papel, ocupam uma área de 793515 ha correspondendo a 23% da área florestal total. O principal objetivo deste trabalho foi a análise da recuperação do padrão sazonal da assimilação bruta de carbono (GPP-Gross primary production) numa parcela experimental de eucaliptal em Pegões (Sul de Portugal), após um corte raso realizado no período de Outubro-Novembro de 2006. A parcela localizava-se num povoamento de eucaliptal de 300 ha, situado na Herdade da Espirra, com uma densidade de 1100 árvores/ha. O povoamento foi explorado em regime intensivo de talhadia sob um ciclo produtivo de 12 anos e caracterizado por um período de 12 meses de atividade fisiológica. Os fluxos de carbono nesta parcela foram medidos durante o período 2002-2010, pelo método de covariância turbulência. Uma seca prolongada em 2004 e 2005, em que a precipitação foi reduzida a valores de 50% da média de longo prazo, e o corte raso, com o início de um novo ciclo produtivo, modificaram a capacidade de sumidouro de carbono do eucaliptal. No período anterior ao corte, o pico máximo de assimilação bruta (2204 g m-2) anual ocorreu em 2002, decrescendo para um mínimo de 1255,1 g m-2 em 2005, no auge da seca. Oito meses após o corte os caules emitidos dos cepos iniciaram um crescimento vigoroso e as novas árvores atingiram uma altura média de 8m em 2010. Em Junho de 2007 o eucaliptal recuperou a sua capacidade de sumidouro de carbono, com uma assimilação bruta de 1621,6 g m-2 em 2010. Os padrões sazonais de assimilação bruta de carbono em 2008 e 2009, com os máximos sazonais ocorrendo entre meados de fevereiro e meados de outubro, foram praticamente opostos aos do período anterior ao corte quando o controlo estomático evitou perdas de água em condições de stress hídrico no período de Verão. A recuperação rápida do jovem eucaliptal foi devida à manutenção do sistema radicular desenvolvido e funcional. Em 2010 o padrão de variação mensal foi análogo ao do período anterior ao corte.
Palavras-chave: Fixação de carbono; controlo estomático; sazonalidade; talhadia; sistema radicular desenvolvido
Résumé
Au Portugal, les peuplements d'eucalyptus (Eucalyptus globulus Labill.) destinés surtout à la production de pâte à papier, couvrent une superficie de 793515 ha correspondant à 23% de la superficie forestière totale. L'objectif principal de cette étude était d'analyser l'évolution saisonnière de la reprise de l'assimilation du carbone brute (GPP-Gross Primary Production) dans une parcelle expérimentale d'eucalyptus à Pegões, (sud du Portugal) après une coupe à blanc menée entre Octobre-Novembre 2006. La parcelle est située dans une plantation d'eucalyptus de 300 ha, à Herdade de Espirra, avec une densité de 1100 arbres / ha. Le peuplement a été exploré de forme intensive en taillis sous un cycle de production de 12 ans et se caractérise par une période de 12 mois d'activité physiologique. Les fluxes de carbone dans cette parcelle ont été mesurés au cours de la période 2002-2010, par la méthode de covariance des turbulences. Une sécheresse prolongée en 2004 et 2005, lorsque la précipitation a été réduite à des valeurs de 50% de la moyenne, et la coupe à blanc, avec le début d'un nouveau cycle, ont modifié la capacité de séquestre du carbone de l'eucalyptus. Dans la période avant la coupe, le pic de l'assimilation brute maximale (2204 gm-2) annuel est survenu en 2002 chutant à un minimum de 1255,1 g-2 en 2005 au plus fort de la sécheresse. Huit mois après la coupe les tiges des souches ont commencé une croissance vigoureuse et les nouveaux arbres ont atteint une hauteur moyenne de 8m en 2010. En Juin 2007, l'eucalyptus a retrouvé sa capacité de séquestre du carbone, avec une assimilation brute de 1621.6 gm-2 en 2010. Les tendances saisonnières de l'assimilation du carbone brut en 2008 et 2009, avec des maximums saisonniers qui se produisent entre la mi-février et mi-octobre étaient presque opposées à ceux de la période antérieure à la coupe quand le contrôle stomatique a empêché des pertes d'eau dans des conditions de stress hydrique en période estivale. La reprise rapide des jeunes l'eucalyptus était due à la maintenance du système racinaire développé et fonctionnel. En 2010, le modèle de variation mensuelle était semblable à la période antérieure à la coupe.
Mots clés: Assimilation du carbone; contrôle stomatique; évolution saisonnière; taillis; système racinaire développé
Introduction
The importance of global warming driven by greenhouse gases, especially carbon dioxide, caused by anthropogenic activities (IPCC, 2007) such as combustion of fossil fuels and deforestation is unquestionable. Climate change is of utmost importance in Mediterranean countries like Portugal with dry and hot summer periods and discrete intense rainy episodes occurring mainly in autumn and winter. In Portugal air temperature increased since 1975 at a rate of 0.5ºC per decade (TOMÉ and MIRANDA, 2004), and eleven of the warmest years between 1860 and 2006 occurred during the 12 years prior to 2006 (VITERBO, 2011).
In Mediterranean climates, with mean annual rainfall of about 700-800 mm, another feature inherent in the context of climate change is the occurrence of anomalies concerning frequency and severity of drought periods. The prolonged drought occurred in 2004 and 2005 in Iberian Peninsula was thereby the most severe of the last 140 years (GARCIA-HERRERA et al., 2007). Climatic scenarios for Portugal based on global scale models point out to air temperature increases of 3ºC to 7ºC and precipitation losses of 20% to 40% till the end of the XXIth century (MIRANDA et al., 2006).
In this context the problem of afforestation with exotic fast growing tree species assumes particular interest. In Portugal, eucalypt (Eucalyptus globulus Labill.) is a exotic forest species introduced in the second half of 1800s (ALVES et al., 2007). Nowadays eucalypt stands are submitted to intensive management aimed primarily for pulp production, with high wood productivity of about 16m3 ha-1 year -1, occupying an area of 739515 ha, corresponding to 23% of total forest area (NATIONAL FOREST INVENTORY, 2005-2006).
Over the past 20 years the scientific knowledge on eddy covariance methodology (AUBINET et al., 2000; REICHSTEIN et al., 2005; PAPALE et al., 2006) and on the factors affecting NEE (Net Ecosystem Exchange), GPP (Gross Primary Production) and TER (Total Ecosystem Respiration)in forests improved substantially. This was due to research projects such as Carboeuroflux (2000–2003), Carboeurope (2004–2008) or scientific networks such as Fluxnet. These factors include weather, latitude, (VALENTINI et al., 2000) or season of the year (FALGE et al., 2002), tree biology, duration of the growing season or temperature and soil moisture (SCHMIDT et al., 2000). In Mediterranean climates, carbon sequestration is decisively influenced by water stress of the ecosystem (REICHSTEIN et al., 2002) and atmospheric flows governed by the dynamics of the functioning of stomata, under conditions of severe summer water stress (GRANIER et al., 2007; PEREIRA et al., 2007; RODRIGUES et al., 2011). Relationships between NEE, GPP and water vapor deficit and photosynthetic active radiation may be obtained e.g. by general estimating equations modeling (RODRIGUES et al., 2011).
Concerning eucalypt species, available evidence from hydrological studies show that besides fast-growing carbon fixation, such species tend to have a very effective stomatal response mechanism to soil moisture deficits and vapor pressure deficit (VPD) that allows trees to survive droughts (NEWSON and CALDER, 1989; PEREIRA et al., 2007). With Eucalyptus globulus Labill., available information points to stomatal responses above VPD thresholds of 1.2kPa (NEWSON and CALDER, 1989; RODRIGUES et al., 2011).
Another general principle, applied to some eucalypt species, is that when the atmospheric evaporative demand is high and trees still have water reserves in the soil, high rates of evapotranspiration can occur (NEWSON and CALDER, 1989). Annual rates of transpiration in Eucalyptus globulus is 2700 mm, an order of four times the annual rainfall of 680mm were listed on the site of Hotham Valley in Western Australia (GREENWOOD et al., 1985). Hydrological studies in eucalypt stands in Portugal and Spain (DAVID et al., 1994, FERNÁNDEZ et al., 2006) also showed that mature and deep root system allows to a fast regeneration of eucalypt stand after a clear felling with substantial increases in evapotranspiration in a short period (3-4 years) after felling. This fast regeneration is linked to significant growth in tree height and LAI (SOARES et al., 2007). The drastic reduction in aerial biomass after felling, simultaneous with an active root system typical of mature trees in their term of production cycle, allow young shoots to withstand a severe summer water stress. One of the main effects of water deficits is precisely to limit leaf growing and canopy expansion (OSÓRIO et al., 1998).
During the period 2002-2010, two main events happened in the eucalypt site with drastic consequences on the ability of carbon assimilation by the ecosystem, namely a prolonged drought in 2004–2005 and a tree felling in October–November 2006.
The effects of two phase drought in carbon fixation were analyzed in another paper (RODRIGUES et al., 2011). In the period 2008-2009 seasonal patterns of GPP, with carbon uptake active from mid-February to mid-October and nearly stopping in winter due to frost, were substantially distinct to the period before the felling, when leaf stomatal closure preventing transpiration cancelled carbon uptake in Summer. In 2010, GPP monthly variation in the young matured stand recovered the pattern prior to the clearcutting.
In this study we focused mainly on the period after felling, by analyzing the recovery of carbon sink capacity concerning the evolution of annual and seasonal patterns of GPP, the tree growth and the evapotranspiration. The main working hypothesis to test was whether our eddy covariance and meteorological data available corroborated available information on Eucalypt globulus Labill, obtained by allometric, physiological and hydrological studies, allowing thereby improvement for coordination of future scientific strategies in addressing such complex and challenging issues.
Material and methods
The experimentation on carbon sequestration in the eucalypt stand in Herdade da Espirra was part of the Carboeuroflux and Carboeurope projects. The site is located in a 300ha eucalypt (Eucalyptus globulus Labill.) stand in the county of Pegões (38º38'N, 8º36'W) in Southern Portugal. The stand is located in a flat area intensively managed as a coppice for pulp production, with a density of 1100 trees/ha and a 12 month growth period. The climate is Mediterranean type with mean annual temperature and precipitation over the long term (1961–1990) of 15.9º C and 709 mm respectively. Under normal conditions the precipitation occurs mainly between October and April, with no rainfall in summer. A tree felling was made in October-November 2006 after a twelve year productive cycle in trees with 20 m average height. The young shoots were submitted to a cultural thinning in October–December 2008, for the removal of three stems of each group of four stems per stump. The new trees showed strong height growth reaching an average 8 m height in December 2010.
Carbon flux and evapotranspiration measurements were made by eddy covariance (AUBINET et al., 2000) at a 21 Hz data sampling rate with an experimental unit installed in 2002 at the top of a 33m watch tower but moved to a 12 m height after the felling. The instrumentation included an ultrasonic Gill, R2 anemometer, an open path IRGA LI-7500 gas analyzer and an automatic weather station (Campbell Scientific CR10 data-logger) for continuously monitoring the main meteorological parameters. Subsequently, after the felling, the eddy covariance unit was moved to a height of 12 m above the ground. The distance from the tower to the edge of the stand varied between 700 m and 1800 m. Calibration of the gas analyzer with a reference gas was carried out annually.
Measurements of eddy fluxes in the constant flux layer were made since January 2002. Data for fluxes consisted in averages over 30 min periods. Half hour fluxes calculation involved two axis coordinate rotation, linear detrending by least squares regression (GASH and CULF, 1996), Webb-Leuning (WEBB et al., 1980) correction for density fluctuations and Schotanus correction for sonic temperature (SCHOTANUS et al., 1983) Instantaneous data with friction velocity below 0.2 ms-1, mean vertical velocity fluctuations higher than 0.35 ms-1, high frequency and low frequency spikes (PAPALE, 2006) were discarded. Gap-filling of missing or bad quality data and a partition of carbon uptake in gross primary production and total respiration (REICHSTEIN et al., 2005) was made with an online algorithm (http://gaia.agraria.unitus.it). An automatic weather station (Campbell Scientific CR10 data logger) was installed to sample meteorological data every 30 s and recording averages over 30 min periods. Precipitation was calculated using the integral of half hour periods data. Mean air temperature was measured at 25.2 m, 26.7 m, 29.2 m and 3.6 m with self produced Cu-Cons thermocouples of 0.15mm diameter. Global incident radiation and net radiation were also measured. For additional details on methodologies of fluxes and meteorological measurements refer to RODRIGUES et al., 2005 and RODRIGUES et al., 2011.
Leaf area index was obtained from MODIS satellite data. Soil carbon storage was evaluated before(2002) and after the felling(2007) by excavation in four points close to watch tower with four replicates at depths 0-25 cm, 25-60 cm and 60-100 cm. Soil samples were oven-dried and total carbon was evaluated by the method described in ISO standard 10694. Measurements of total tree height, crown length and diameter at breast height were made in January 2002, 2003, 2005, and 2006 in five plots of 225 m2 adjacent to the tower, for allometric estimation of total carbon biomass. The equations used were these reported by ANTÓNIO et al. (2007) for individual trees.
The values of soil moisture and temperature were measured continuously every two hours since January 2007 onwards, at depths of 10 cm, 20 cm, 30 cm, 40 cm, 60 cm and 1 m with a Delta-T probe, model PR2.
Results and discussion
Meteorological variables
The annual results of accumulated NEE, GPP, TER, precipitation (Prec), E (mm) and global solar radiation (Rg) and mean air temperature (Ta), are shown in Table 1. The average temperatures in 2003, 2004, 2005, 2006, 2009 and 2010 exceeded the long-term average of 15.9ºC. The annual rainfall in 2004 and 2005 was reduced by 47% compared to the long-term average of 709 mm. (Figure 1). Throughout the period covered by this study, accumulated rainfall during June–September was less than 9% of total precipitation. The drought years of 2004 and 2005, besides the lower annual rainfall, showed a marked asymmetry in the distribution of rainfall throughout the year. Indeed the precipitation occurred in the first quarter of 2004 accounted for 55.5% of total annual and in 2005 the accumulated rainfall in the last quarter represented 40% of the total annual. Precipitation in 2008 and 2009 was more uniform.
Table 1 - Cumulative totals of NEE GPP and TER, mean air temperature (Ta), global solar radiation, (Rg), rainfall (Prec), and evapotranspiration (E)
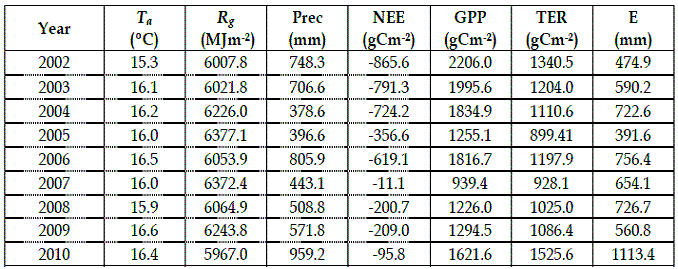
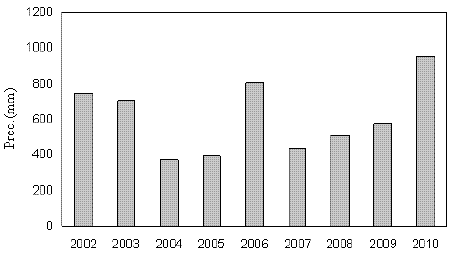
Figure 1- Evolution of annual precipitation in the whole period
The highest values of average monthly atmospheric vapor pressure deficit (VPD) in the two drought years were 7.56 hPa and 8.67 hPa respectively. Monthly average VPD decreased to 5.34 hPa in the period after tree cutting. Precipitation after the felling increased to 443.1 mm, 508.8 mm, 571.8 mm and 959.2 mm, in 2007, 2008, 2009 and 2010, respectively. Over the whole years, incident solar radiation and temperature showed a relatively uniform variation, not comparable to seasonal and annual variations of precipitation in dry years. In 2007–2010 soil moisture increased annually with depth between 3.47% at 10 cm and 11.49% at 1 m, surpassing wilting point in sandy soils (5%) below 60 cm. Soil moisture depth showed also average monthly variation between 5% in August–October and 11% in November-February.
Carbon fluxes and Biomass
The impact on GPP derived from the drastic reduction of precipitation in 2004 and 2005 was felt mainly in 2005. Indeed, the values of GPP were respectively 2206.0 gCm-2, 1995.6 gCm-2, 1834.9 gCm-2 (Table 1 and Figure 2) in 2002, 2003 and 2004 respectively, dropping to 1255.1 gCm-2 in 2005 (Table 1). In 2004, the first drought year, the annual rainfall of 378.6 mm was of a half order of the total evapotranspiration of 722.6 mm, depleting soil water and sustaining values of GPP to levels of the same order of 2002 and 2003. This agrees with the general principle applied to some eucalypt species that is, when atmospheric evaporative demand is high and trees still have water reserves in the soil, high rates of evapotranspiration can occur (NEWSON and CALDER, 1989; GREENWOOD et al., 1985). In the period prior to felling the average monthly NEE and GPP were maximum in the middle spring and minimum in late summer (Figure 3). Its variation was opposite to VPD, with the September fall in carbon uptake coinciding with maximum monthly VPD. This is due to the fact that forest stomatal conductance tends to be higher at low VPDs, as shown, e.g., by DAVID et al. (1997) for an eucalypt stand in Portugal. The values of leaf area index (LAI) obtained by remote sensing from MODIS satellite varied seasonally around 5 in the period before the felling. Averaged monthly LAI decreased in 2005 due to leaf yellowing associated with intense water stress.
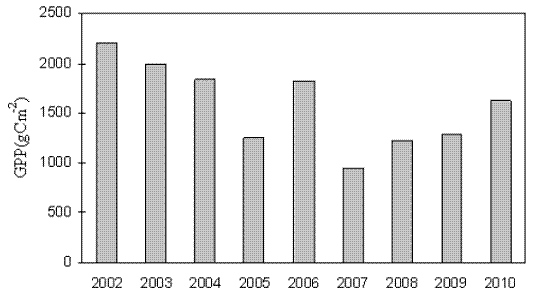
Figure 2 - Annual GPP for the whole period

Figure 3 - Monthly average GPP (+ avg. 2003-2006, ---2008, ◊ 2009, — 2010)
After the felling in October–November 2006 the eucalypt coppice, deprived of foliage, become a carbon source. New shoots emerged from the remaining stumps and the stand recovered its carbon sink capacity after June 2007, with annual values of GPP of 1226 gCm-2, 1294.5 gCm-2, and 1621.6 gCm-2 in 2008, 2009 and 2010, respectively (Table 1). Typical day curves of GPP on a quarterly basis in averaged periods 2002–2003 and 2009–2010, reflect the tendencies of monthly GPP. Indeed, Fig. 4B shows that in the averaged period 2009–2010, mean daily GPP was higher in July-September than in winter. Oppositely, in the averaged period 2002–2003 (Figure 4A) GPP values were higher in all quarters and in winter GPP was higher than in summer. On the other hand in 2002–2003, as a consequence of the effects of the summer water stress effect in the mature stand, GPP showed an asymmetrical pattern at noon much more obvious than in 2009–2010 (Figure 4).
Figure 4 - Typical day curves for average GPP: (A) avr. 2002-2003, (B) avr (2009-2010) (+ Jan.-Mar. ---Apr.-Jun. ♦ Jul.-Sep. ▬ Oct.-Dec.)
Typical day curves in the whole 2002-2010 period (not shown) indicated that as rule global solar radiation phased and peaked with NEE and GPP at about noon and TER phased with VPD and air temperature (RODRIGUES et al., 2011). Typical day curves of evapotranspiration and GPP were also synchronized showing the fundamental role of leaf stomata on the linking of carbon and water fluxes.
The annual averaged values of MODIS LAI of young plants increased to 2.4, 3.4, 2.8 and 3.1 in 2007, 2008, 2009 and 2010 respectively, reflecting a lower aerial biomass, comparatively to the period before the felling. Mean LAI in 2008 was reduced by the thinning in the last quarter. Anyway this significant LAI increase is associated to a fast growth in the young plants up an average 8m height at the end of 2010.The young plants were sensitive to the effects of winter frost in the four years after the cutting, with crisp leaf yellowing processes and diminishing of LAI and GPP.
The fast regeneration of the eucalypt stand agrees with allometric data of SOARES et al., (2007) and with the aforementioned conclusions of hydrological studies about the maintenance of a mature and viable root system (DAVID et al., 1994; FERNÁNDEZ et al., 2006) able to extract water at deeper levels with higher soil moisture. Thereby the new plants with a reduced aerial biomass and a developed root system were able to withstand summer water stress and assimilating carbon in summer under higher solar radiation. The monthly pattern of GPP in 2008 and 2009 (Figure 3) was thus changed, almost reversed peaking in summer when higher solar radiation is available, without the leaf stomata control typical of mature trees with higher aerial biomass under high water stress. On the other hand, GPP in these years decreased in winter, especially in January, due to the thinning in October-December 2008 and to a higher sensibility to frost. Water stress during summer periods after the felling was also minimized by a more uniform precipitation in 2008 and 2009, simultaneous with lower monthly VPD. In 2010, despite a decrease in winter, monthly GPP peaked at late spring (Figure 3) almost coinciding with the corresponding maximum in 2002–2003. Thereby it is expected that the pattern change of seasonality should be temporary and abolished as the aerial biomass of the new trees increases. A four year transient period associated to a recovery of fast full year leaf production and aerial biomass and an increase in evapotranspiration, was also reported in eucalypt plots after felling by SAMRAJ et al., 1988 and SHARDA et al., 1998 in India and by DAVID et al., 1994 and SOARES et al., 2007 in Portugal and FERNANDEZ et al., 2006 in Spain. DAVID et al., 2007 also report that the intensive management in terms of fertilization and soil tillage, as happened in Espirra plot, and high water use efficiency are determinant factors contributing to this fast growth by eucalypt coppices after felling.
An average total biomass of 123 ton ha-1 (or 61.5 ton C ha-1) was estimated at the end of rotation, with an annual increment of about 19 ton year-1 in 2002-2003, dropping to 7.8 ton year-1 in the dry period 2005-2006. This amount of biomass accumulated in the twelve year productive cycle is consistent with the average annual NEE values or the order of 700-800 gCm-2, indicated in Table 1. After the tree felling in an area of 200 ha, an average 13.6 ton biomass ha-1 oflitter (leaves, branches and twigs) was left in the soil (RODRIGUES et al., 2011). Belowground carbon was estimated as 38.2t/ha in the soil till 1m depth, corresponding to 15 t/ha, 12.4 t/ha and 10.8 t/ha in the 0-25 cm, 25-60 cm and 60-100 cm soil layers, respectively.
Conclusions
The eucalyptus coppice submitted to intensive management was a substantial carbon sink with average NEE and GPP of about -790 gCm-2 and 2000 gCm-2 in the period 2002–2004, when the effects of lack of precipitation were not felt. GPP diminished after the felling and GPP monthly pattern also changed, especially in 2008–2009 with higher carbon assimilation in summer, oppositely to the period before felling when summer carbon uptake practically stopped.
The results of eddy covariance methodology reinforced the hypothesis, made firstly under hydrological and physiological studies, whereby the maintenance of a mature root system in a coppiced eucalypt stand is crucial to guarantee a fast growing of young plants. Indeed, not only GPP increased significantly during the four year period after felling, accompanying the strong tree height growth, but also carbon assimilation in summer was possible in 2008 and 2009, given the imbalance between aerial and root biomass. This profile tendency proofed transient till 2010, when supposedly trees begun to achieve maturity and the stand began to invert to the situation prior to the felling.
References
ALVES, A.M., PEREIRA, J.S., SILVA, M.N., 2007. A introdução e a expansão do eucalipto em Portugal. In O eucaliptal em Portugal, Impactes Ambientais e Investigação Científica. The beginning and expansion of eucalypt in Portugal. Edts.: Alves, A.M., Pereira S.J., and Silva J.M.N., ISAPress, pp. 13-24.
ANTÓNIO, N., TOMÉ, M., SOARES, P., TOMÉ, J., FONTES, A.L., 2007. Effect of stand and site variables on the estimation of above ground tree biomass of Eucalyptus globulus. Canadian Journal of Forest Research 37: 895-906. [ Links ]
AUBINET, M., GRELLE, A., IBROM, A., RANNIK, Ü., MONCRIEFF, J., FOKEN, T., KOWALSKI AS, MARTIN PH, BERBIGIER, P., BERNHOF, CH, CLEMENT, R., ELBERS, JA, GRANIER, A., GRÜNWALD, T., MORGENSTERN, K., PILLEGAARD, K., REBMANN, C., SNIJDERS, W., VALENTINI, A., VESALA, T., 2000. Estimates of the annual net carbon and water exchange of forests: Euroflux methodology. Advances in Ecological Research 30: 113-175. [ Links ]
DAVID, J.S., HENRIQUES, M.O., DAVID, T.S., TOMÉ, J., LEDGER, D.C., 1994. Clearcutting effects on streamflow in coppiced Eucalyptus globulus stands in Portugal. Journal of Hydrology 162: 143-154. [ Links ]
DAVID, T.S., FERREIRA, M.I., DAVID, J.S., PEREIRA, J.S., 1997. Transpiration from a mature Eucalyptus globulus plantation in Portugal during a Spring–Summer period of progressively higher water deficit. Oecologia 110: 153–158. [ Links ]
FALGE, E., BALDOCCHI, D.D., TENHUNEN, J., AUBINET, M., BAKWIN, P., BERBIGIER, P., BERNHOF, C., BURBA, G., CLEMENT R., DAVIS, K.J., ELBERS, M. GOLDSTEIN, A.H., GRELLE, A., GRANIER, A., GUDMUNDSSON, J., HOLLINGER, D., KOWALSKI, AS, KATUL, G., LAW, B.E., MALHI, Y., MEYERS, T., MONSON, R.K., MUNGER, J.W., OECHEL, W., PAW, U., PILEGAARD, K.T., RANNIK, K., REBMANN, U., SUYKER, C., THORGEIRSSON, A., VALENTINI, H., WILSON, R., WOFSY, K.S., 2002. Seasonality of ecosystem respiration and gross primary derived from the production FLUXNET measurements. Agricultural and Forest Meteorology 113: 53-74. [ Links ]
FERNÁNDEZ, C., VEGA, J.A., GRAS, L.M., FONTURBEL, T., 2006. Changes in water yield after a sequence of perturbations and forest management practices in an Eucalyptus globulus Labill. watershed in Northern Spain. Forest Ecology and Management 234: 275-281. [ Links ]
IPCC, 2001. Climate Change 2001: The Scientific Basis, Contribution of the Working Group I to the Third Assessment Report of the Intergovernmental Panel of Climate Change. Cambridge University Press. [ Links ]
IPCC, 2007.Climate Change 2007: The Scientific Basis-Summary for Policymakers. Contribution of the Working Group I to the Fourth Assessment Report of the Intergovernmental Panel of Climate Change. Cambridge University Press. [ Links ]
ISO 10694: 1995. Soil quality - Determination of organic and total carbon after dry combustion (elementary analysis). International Organization for Standardization, 01 Mar 1995, 7 pp. [ Links ]
GARCIA-HERRERA, R., PAREDES, D., TRIGO, R.M., TRIGO, I.F., HERNANDEZ, E., BARRIOPEDRO, D., MENDES, M.A., 2007. The outstanding 2004/05 drought in the Iberian Peninsula: Associated atmospheric circulation. Journal of Hydrometeorology 8: 483-498. [ Links ]
GASH, J.H., CULF, A., 1996. Applying a Linear Detrend to Eddy Correlation Data in Real Time. Boundary Layer Meteorology 79: 301-306. [ Links ]
GRANIER A., REICHSTEIN, M., BREDA, N., JANSSENS, IA, FALG, E., CIAL, P., GRUNWALD, T., AUBINET, M., BERBIGIER, P., BERNHOF, C., BUCHMANN, N., FACINAS, O., GRASSI, G., HEINESCH, B., ILVESNIEMI, H., KERONEN, P., KNOHL, A., KOSTNER, B., LAGERGREN F., LINDROTH, A., LONGDOZ, B., LOUSTAU, D., MATTHEW J., MONTAGNANI, L., NYS, C., MOORS, E., PAPALE, D., PEIFFER, M., PILEGAARD K., PITA, G., PUMPANEN, J., RAMBAL, S., REBMANN, C., RODRIGUES A., SEUFERT, G., TENHUNEN J., VESALA, T., WANG, Q., 2007. Evidence for soil water control on carbon and water dynamics in European forests During the extremely dry year: 2003. Agricultural and Forest Meteorology 143: 123-145. [ Links ]
GREENWOOD, E.A.N., KLEIN L., BERESFORD, J.D., WATSON, G.D., 1985. Differences in annual evaporation between grazed pasture and Eucalyptus species in plantation on a saline farm catchment. Journal of Hydrology 78: 357-366. [ Links ]
JANSSENS, I.A., LANKREIJER, H., MATTEUCCI, G., KOWALSKI AS, BUCHMANN, N., EPRON, D., PILEGAARD, K., KUTSCH, W., LONGDOZ, B., GRÜNWALD, T., MONTAGNANI L., DORE, S., REBMANN, C., MOORS, E.J., GRELLE, A., RANNIK, Ü., MORGENSTERN, K., CLEMENT, R., GUDMUNDSSON, J., MINERBO, S., BERBIGIER, P., IBROM, A., MONCRIEFF, J., AUBINET, M., BERNHOF, C., JENSEN, N.O., VESALA, T., GRANIER, A., SCHULZE, E.D., LINDROTH, A., DOLMAN, A.J., JARVIS, P.G., CEULEMANS, R., VALENTINI, R., 2001. Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Global Change Biology 7: 269-278. [ Links ]
MIRANDA, P., VALENTE, A., TOMÉ, A., TRIGO, R., COELHO, M.F., AGUIAR, A., AZEVEDO, E., 2006. O clima de Portugal nos séculos XX e XXI. Alterações climáticas em Portugal- Cenários, Impactes e medidas de adaptação. Ed. F.D. Santos e P. Miranda, pp. 47-113. Lisboa, Gradiva. [ Links ]
NEWSON, M.D., CALDER, I.R., 1989. Forests and Water resources in Proceedings of a Royal Society Discussion Meeting, held on 2- 3 June, pub. Philosophical Transactions of the Royal Society of London, University Press, Cambridge, pp. 283-298. [ Links ]
OSÓRIO, J., OSÓRIO, M.L., CHAVES, M.M., PEREIRA, J.S., 1998. Effects of water deficits on 13C discrimination and transpiration efficiency of Eucalyptus globulus clones. Australian Journal of Plant Physiology 25: 645-653. [ Links ]
PAPALE, D., REICHSTEIN, M., AUBINET, M., CANFORA, E., BERNHOFER, C., KUTSCH, W., LONGDOZ, B., RAMBAL, S., VALENTINI, R., VESALA, T., YAKIR, D., 2006. Towards a standardized processing of net ecosystem exchange measured with eddy covariance technique: algorithms and uncertainty estimation. Biogeosciences 3: 571-583. [ Links ]
PEREIRA, J.S., MATTHEW, J.A., AIRES, L.M., PITA, G., PIO, C. DAVID, J.S., ANDRADE, V., BANZA, J. DAVID, T.S., PAÇO, T.A., ROBERTS, A., 2007. Net ecosystem carbon exchange in three contrasting Mediterranean ecosystems - the effect of drought. Biogeosciences 4: 791-802. [ Links ]
REICHSTEIN, M., TENHUNEN, J.D., ROUPSARD, O., OURCIVAL, J.M., RAMBAL, S., MIGLIETTA, F., PERESSOTTI, A., PECCHIARI, M., TIRONE G., VALENTINI, R., 2002. Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: revision of current hypotheses? Global Change Biology 8(10): 999-1017. [ Links ]
REICHSTEIN, M., FALGE, E., BALDOCCHI, D., PAPALE, D., AUBINET, M., BERBIGIER, P., BERNHOF, C., BUCHMANN, N., GILMANOV, T., GRANIER, A., GRUNWALD, T., HAVRANKOVA, K., ILVESNIEMI, H., JANOUS, D., KNOHL, A., LAURILA, T., LOHILA, A., LOUSTAU, D., MATTEUCCI, G., MEYERS, T., MIGLIETTA, F., OURCIVAL, JM, PUMPANEN, J., RAMBAL, S., ROTENBERG, E., SANZ, M., TENHUNEN, J., SEUFERT, G., VACCARI F., VESALA, T., YAKIR , D., VALENTINI, R., 2005. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm. Global Change Biology 11(9): 1424-1439. [ Links ]
RODRIGUES, A., PITA, G., MATEUS, J., 2005. Turbulent fluxes of carbon dioxide and water vapor over the eucalyptus forest in Portugal. Silva Lusitana 13(2) 169-180. [ Links ]
RODRIGUES, A., OLIVEIRA, H., 2006. Sequestro de carbono: Tendências globais e perspectivas da floresta portuguesa. Ingenium 92: 68-71. [ Links ]
RODRIGUES, A., PITA, G., MATEUS J., KURZ-BESSON, C., CASQUILHO, M., CERASOLI, S., GOMES, A., PEREIRA, J., 2011. Eight years of continuous carbon fluxes measurements in Portuguese eucalypt stand under the two main events: Drought and felling. Agricultural and Forest Meteorology 151: 493-507. [ Links ]
SOARES, P., TOMÉ, M., PEREIRA, J.S., 2007: A produtividade do eucaliptal in O eucaliptal em Portugal: Impactes ambientais e investigação científica Edts.: Alves, A.M., Pereira S.J., and Silva J.M.N., ISAPress, pp. 27-61. [ Links ]
SAMRAJ, P., SHARDA, V.N., CHINNAMANI, S., LAKSMAN, V., HALDORAI, B., 1998. Hydrological behavior of Nilgiri subwatersheds as affected by bluegum plantations. Part I- The annual water balance. Journal of Hydrology 103: 335-345. [ Links ]
SHARDA, V.N., SAMRAJ, P., SAMRA, J.S., LAKSMANAN, V., 1998. Hydrological behavior of first generation bluegum plantantions in the Nilgiri subwatersheds. Journal of Hydrology 211: 50-60. [ Links ]
SCHMID, H.P., GRIMMOND, C.S.B., CROPLEY, F., OFFERLE, B., SU, H.B., 2000. Measurements of CO2 and energy fluxes over a mixed hardwood forest in the mid-western United States. Agricultural and Forest Meteorology 103: 357-374. [ Links ]
SCHOTANUS, E.K., NIEUWSTADT, F.T.M., DE BRUIN, H.A.R., 1983. Temperature measurement with a sonic anemometer and its application to heat and moisture flux. Boundary-Layer Meteorology 26: 81-93. [ Links ]
TOMÉ, A.R., MIRANDA, P.M.A., 2004. Piecewise linear fitting and trend changing points of climate parameters. Geophysical Research Letters, 31, L02207, doi:10.1029/ 2003G019100. [ Links ]
VALENTINI, R., MATTEUCCI, G., DOLMAN, A.J., SCHULZE, E.D., REBMANN, C., MOORS, E.J., GRANIER, A., GROSS, P., JENSEN, N.O., PILEGAARD, K., LINDROTH, A., GRELLE, A., BERNHOFER,C., GRUNWALD, T., AUBINET, M., CEULEMANS, R., KOWALSKI, A.S., VESALA, T., RANNIK,U., BERBIGIER, P., LOUSTAU, D., GUDMUNDSSON, J., THORGEIRSSON, H., MONTAGNANI, L.,MINERBI, S., JARVIS, P.G., 2000. Respiration as the main determinant of carbon balance in European forests. Nature 404: 861-865. [ Links ]
VITERBO, P., ESPÍRITO SANTO, F., BERNARDINO M., 2011. Climate change observed and climate scenarios in Portugal. Presentation made to the "Workshop of adaptation of agriculture to climate change", April, 19th L-INIA, Oeiras, Portugal. [ Links ]
WEBB, E.K., PEARMAN, G.I. and LEUNING, R., 1980. Correction of Flux Measurements for Density Effects Due to Heat and Water Vapor Transfer. Quarterly Journal of the Royal Meteorological Society 106: 85-100. [ Links ]
Entregue para publicação em outubro de 2012
Aceite para publicação em dezembro de 2012
