










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkSilva Lusitana
versão impressa ISSN 0870-6352
Silva Lus. vol.19 no.1 Lisboa 2011
Resposta Fisiológica de Pinus spp. nas Primeiras Horas após Infecção com Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae)
Carla Santos* e Marta Vasconcelos**
*Bolseira de Investigação
**Investigadora Auxiliar
CBQF – Escola de Biotecnologia. Universidade Católica Portuguesa. Rua Dr. António Bernardino de Almeida, 4200-072 PORTO
Sumário
O objectivo deste estudo consistiu na comparação da resposta à infecção com o nemátode da madeira do pinheiro (NMP) Bursaphelenchus xylophilus entre Pinus pinaster e Pinus pinea no estádio inicial da doença, três horas após inoculação. O NMP é o agente causal da doença da madeira do pinheiro, tem em Portugal como hospedeiro principal o pinheiro bravo, P. pinaster, e tem como vector Monochamus galloprovincialis.
Actualmente, não há estudos que descrevam os efeitos da infecção do nemátode nas plantas a um estádio inicial da doença e, no que diz respeito aos efeitos fisiológicos e metabólicos nas plantas, verificou-se uma diminuição de 10,9% no teor de água em P. pinaster (por comparação com árvores inoculadas com água) e 6,7% em P. pinea; relativamente à concentração de clorofilas, uma diminuição para cerca de metade do valor controlo foi registado para ambas as espécies, mas mais pronunciadamente em P. pinaster. Um estudo exploratório com cromatografia gasosa também foi efectuado, que demonstrou que os compostos voláteis produzidos permitem uma clara diferenciação entre espécies, mas não entre inoculação controlo vs. nemátode.
Palavras-chave: Concentração de clorofilas; conteúdo de água; cromatografia gasosa
Physiological Response of Pinus spp. In the First Hours After Infection with Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae)
Abstract
The purpose of this study was to compare the response to infection with the pine wood nematode (PWN) Bursaphelenchus xylophilus between Pinus pinaster and Pinus pinea at an initial stage of the disease, three hours after inoculation. The PWN is the causal agent of pine wilt disease, in Portugal its main host is the maritime pine, P. pinaster, and its vector is Monochamus galloprovincialis.
Currently there are no studies that describe the effects of nematode infection in plants at an early stage of the disease and with regards to the physiological and metabolic indicators of the disease response in the plants, there was a decrease of 10.9% of water content in P. pinaster (compared with trees inoculated with water) and 6.7% in P. pinea; in the study of total chlorophyll concentration, a decrease to about half of the control value was recorded for both species, but more pronounced in P. pinaster.
An exploratory study was also made with gas chromatography, which showed that the volatile compounds produced were distinct between species, but not between control vs. nematode-inoculated plants.
Key words: Chlorophyll concentration; water content; gas chromatography
Réponse Physiologique de Pinus spp. aux Premières Heures aprés l'Infection par Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae)
Résumé
Le but de cette étude était de comparer la réponse à l'infection par le nématode du pin (NDP) Bursaphelenchus xylophilus entre Pinus pinaster et Pinus pinea dans la phase initiale de la maladie, trois heures après l'inoculation. Le NDP est l'agent causal de la maladie de dépérissement du pin, au Portugal son hôte principal est le pin maritime, P. pinaster, et son vecteur Monochamus galloprovincialis.
Actuellement, il n'existe pas d'études qui décrivent les effets de l'infection par le nématode des plantes à un stade précoce de la maladie et en ce qui concerne les indicateurs physiologiques et métaboliques chez les plantes, il ya eu une diminution de la teneur en eau de 10,9% en P. pinaster (comparaison avec les arbres inoculés avec de l'eau) et 6,7% en P. pinea; dans l'étude de la concentration de chlorophylle totale, une diminution d'environ la moitié de la valeur de commande a été enregistrée pour les deux espèces, mais plus prononcée dans P. pinaster.
Une étude exploratoire a également été faite avec chromatographie en phase gazeuse, qui a montré que les composés volatils produits permettent une nette différenciation entre les espèces, mais pas entre le contrôle des plantes contre les nématodes inoculés.
Mots clés: Concentration de chlorophylle; teneur en eau; chromatographie en phase gazeuse
Introdução
O nemátode da madeira do pinheiro (NMP), Bursaphelenchus xylophilus (Steiner and Buhrer, 1934) Nickle, 1970 é o agente causal da doença da madeira do pinheiro (DMP) (NICKLE et al., 1981). O género, Bursaphelenchus Fuchs 1937 inclui quase 90 espécies e faz parte da Família Aphelenchoididae. Os nemátodes que pertencem a este género podem ser micofágicos, parasitas de plantas, ou ambos, sendo que B. xylophilus pertence à última categoria (ZHAO et al., 2008).
Esta doença mata, tipicamente, pinheiros com mais de 10 anos de idade em períodos de tempo que podem ser entre poucas semanas a meses e, apesar de todos os avanços alcançados nesta área, o mecanismo da doença ainda não foi clarificado (ZHAO et al., 2008). No seu país nativo, EUA, o nemátode não é considerado um agente patogénico, mas após a sua exportação, as consequências foram massivas nas florestas mundiais, o que levou a um abalo grave na economia de cada zona afectada (ZHAO et al., 2007).
Em Portugal, o NMP é transportado pelo vector Monochamus galloprovinciallis, uma espécie endémica (NAVES et al., 2007), e o seu hospedeiro principal é P. pinaster Ait. (MOTA et al., 1999). O pinheiro bravo é muito importante para a economia portuguesa, na medida em que ocupa mais de um milhão de hectares das florestas nacionais e vários estudos têm vindo a ser direccionados para o seu melhoramento genético (AGUIAR et al. 2003; CORREIA et al. 2004). As espécies Pinus sylvestris e Pinus halepensis também são possíveis hospedeiros do NMP, porém a sua distribuição e abundância é limitada em Portugal (MOTA e VIEIRA, 2008). Estudos recentes, sob condições laboratoriais, demonstraram que as populações portuguesas de M. galloprovincialis demonstram preferência por P. sylvestris em relação a outras espécies de pinheiro (KOUTROUMPA et al., 2009). Estas preferências alternativas podem eventualmente redireccionar o nemátode, B. xylophilus, a infestar outros tipos de árvores, e é por isto que são necessários mais estudos, dado que a flora portuguesa é tão diversa.
P. pinea é outra espécie de pinheiro importante em Portugal, dado que, na natureza, não é afectada pela doença. Aparentemente, o vector não se alimenta, nem coloniza estes pinheiros; no entanto, o nemátode é capaz de invadir, multiplicar-se, infectar e matar esta árvore (ZHAO et al., 2008), apesar de mais lentamente do que em P. pinaster (MOTA e VIEIRA, 2008).
Tendo em conta os riscos de dispersão da doença, foram desenvolvidos regulamentos rigorosos para eliminar a peste, particularmente aquando da importação de madeira e derivados (DWINELL, 1997). Após a detecção do nemátode em pinheiro bravo português, foi desenvolvido e implementado o programa de controlo "PROLUNP".
No que respeita à detecção visual da doença, durante os diferentes estádios de infecção (desde o inicial ao avançado) os sintomas vão-se agravando e acabam por resultar na morte do hospedeiro. Entre os principais sintomas encontra-se a cessação de exsudação de resina nos caules, diminuição da fotossíntese, degenerescência das células parenquimatosas e xilémicas, formação traumática de canais de resina, produção de substâncias fitotóxicas, aumento da taxa respiratória e da síntese de etileno (FUKUDA, 1997). Para além destes, o sintoma mais crítico é o amarelecimento das agulhas, que aparece no estádio mais tardio da infecção, até que a árvore fica castanha por completo, o que acaba por complicar a detecção atempada da doença (GLEASON et al., 2000).
De acordo com RUTHERFORD et al. (1992), B. xylophilus move-se mais rapidamente quando as temperaturas são elevadas, o que explica o desenvolvimento drástico de sintomas durante o verão. Nestas condições, a árvore sofre stress hídrico e a fotossíntese nas agulhas diminui, o que acelera o desenvolvimento dos sintomas e conduz à morte das árvores infectadas (FUKUDA, 1997). Esta diminuição do conteúdo de água, quer nos caules, quer nas agulhas, tem vindo a ser estudada e ocorre usualmente pouco antes dos sintomas de infecção serem visíveis em árvores inoculadas (TAN et al., 2005). Deste modo, o teor de água e a sua retenção é considerado um factor de grande peso na patologia da doença do nemátode da madeira do pinheiro (BOLLA et al., 1986) e o stress causado pelas temperaturas elevadas é também um factor abiótico muito preponderante na aceleração da morte do pinheiro, dado que promove a multiplicação de B. xylophilus (SIKORA e MALEK, 1991).
Os monoterpenos são um grupo de voláteis resultantes do metabolismo secundário das árvores e têm um papel muito importante na mediação das interacções entre planta e ambiente, podendo induzir respostas de defesa, que ajudam as plantas a diferenciar entre danos causados por insectos e danos mecânicos (MATEUS et al., 2010). Determinados voláteis são comummente activados nas vias biosintéticas de várias plantas, como o a-pineno (que estima-se fazer parte em 85% dos óleos essenciais das pináceas), b-pineno, limoneno e mirceno (PROENÇA DA CUNHA, 2005) e, quando infectadas, estas árvores emitem um conjunto característico de compostos voláteis que atraem o vector Monochamus. Os óleos essenciais também têm vindo a ser estudados como potenciais agentes de controlo contra a doença, dado que alguns possuem actividade nematicida (KIM et al., 2008).
A cromatografia gasosa – espectrofotometria de massa (GC-MS) é o método mais frequentemente usado na identificação de voláteis, mas a sua aplicabilidade é limitada, dado que os voláteis emitidos pelas árvores são altamente complexos, o que dificulta a separação de todos os componentes (MATEUS et al., 2010). Na maioria dos estudos, os voláteis têm vindo a ser isolados por destilação-extracção simultânea (SDE), que inclui um solvente de extracção, que frequentemente é co-eluído com picos menos retidos numa corrida de cromatografia (SANTOS et al., 2006). Em alternativa, o método de microextracção em fase gasosa de headspace (HS-SPME) tem vindo a ser implementado, dado que dispensa a intervenção de solventes (Supleco, Bulletin 923A).
Crê-se que, à medida que a invasão do nemátode começa, uma resposta hipersensitiva é activada, que resulta na libertação de fenólicos, síntese de toxinas e fitoalexinas e na compartimentalização do xilema e outros tecidos, seguido da inundação dos traqueídeos com oleoresina e substâncias tóxicas (MYERS, 1988). Estudos recentes demonstram que estes mecanismos de defesa contra agentes patogénicos ocorrem nas primeiras horas após infecção (BALDO et al., 2010).
Neste trabalho foram realizados estudos sobre diferenças fenotípicas e metabólicas entre pinheiro infectado e não-infectado das espécies P. pinea e P. pinaster, a um ponto inicial da infecção, dado que é considerado que imediatamente nas primeiras horas após infecção é activada uma resposta hipersensitiva de defesa nas plantas.
Materiais e métodos
Material vegetal
As plantas utilizadas nestes estudos foram Pinus pinea e Pinus pinaster com aproximadamente 1 ano de idade (com alturas de 50±5 cm), provenientes do Viveiro Anadiplanta (Anadia, Aveiro; 40º26'27''N, 8º25'47''W). As árvores foram transferidas dos recipientes para vasos com partes iguais de turfa negra (Siro Plant) e vermiculite exfoliada e foram crescidas em ambiente controlado, numa câmara fitoclima Aralab 10000EHF, com humidade relativa de 80% e um fotoperíodo de 16h dia / 8h noite, com cerca de 490 mmol/m2s de intensidade luminosa. A temperatura foi mantida a 24-26ºC durante o período de dia e a 19-20ºC durante o período da noite.
Cultura de nemátodes
Pequenos quadrados com cerca de 1cm2 de Potato Dextrose Agar com Botrytis cinerea, crescido a 26ºC por 7 dias, foram transferidos para 5 g de grãos de cevada Seara®, previamente autoclavados em tubos de ensaio com 10 ml de água desionizada. Estes tubos foram incubados nas mesmas condições anteriormente descritas e seguidamente, pequenos pedaços de cevada com B. xylophilus isolado geográfico HF (isolado da Região de Setúbal) crescido previamente, foram colocados dentro dos tubos onde o fungo havia sido crescido. Os nemátodes multiplicados foram extraídos pela técnica dos funis de Baermann (BAERMANN, 1917) antes da inoculação e, apenas nemátodes extraídos dentro de um período de 2h é que foram utilizados nas experiências subsequentes.
Inoculação e tempo de amostragem
Nos testes realizados, foram estabelecidos quatro tratamentos diferentes, nomeadamente, árvores P. pinaster e árvores P. pinea selecionadas aleatoriamente e inoculadas com B. xylophilus estirpe HF e árvores P. pinaster e árvores P. pinea selecionadas aleatoriamente e inoculadas com água desionizada (controlo). Para a determinação do conteúdo de água e extracção e quantificação de clorofilas totais, foram utilizadas 30 réplicas biológicas; para a análise dos compostos voláteis foram utilizadas 7 réplicas biológicas.
De acordo com a técnica descrita por FUTAI e FURUNO (1979), uma ferida de 3-5 cm foi feita no caule da árvore com um bisturi a cerca de 40 cm acima do nível do solo. Uma suspensão de 1000 nemátodes em 650 ml de água desionizada foi inoculada nas feridas que, seguidamente, foram cobertas com parafilme para evitar a secagem do inóculo e saída dos nemátodes. O mesmo procedimento foi seguido para as árvores controlo, utilizando 650 ml de água desionizada.
Três horas após inoculação (3 hai), para cada amostra experimental, o caule completo foi recolhido, cortado em pequenos pedaços e armazenado a -80ºC até realização das análises futuras.
Determinação do conteúdo de água
Os caules de cada tratamento foram liofilizados por 72 h; o conteúdo em água e a diminuição do conteúdo de água foram calculados como demonstrado por TAN et al. (2005), pelas seguintes equações:
Conteúdo de água (%) = (peso da amostra fresca – peso da amostra liofilizada) x 100 / peso da amostra fresca;
Diminuição do conteúdo de água (%) = (conteúdo de água da planta controlo – conteúdo de água da planta inoculada) x100 / conteúdo de água da plana controlo.
Extracção e quantificação das clorofilas totais
Seguiu-se o protocolo de ABADÍA et al. (1984), com algumas modificações. Foram adicionados 12,5 ml duma solução de CaCO3 em metanol (4 g/L) a 0,5 g de agulhas de pinheiro de cada amostra individual. Após um período de incubação de 48 h no escuro, à temperatura ambiente, 1 ml da solução metanólica foi diluído em 25 ml de água desionizada e a absorvância foi lida a 663 nm e 645 nm. A quantificação espectrofotométrica de clorofilas foi calculada segundo a equação:
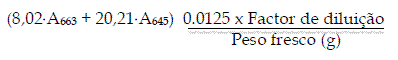
Análise de compostos voláteis
Foi realizada uma análise exploratória do padrão dos compostos voláteis com um cromatógrafo de gás HP 5890A (Hewlett Packard, USA) equipado com um detector de ionização por chama (GC-FID). A separação foi realizada numa coluna Stabilwax-DA com 60m, 0.25mmID, 0.25µm (RESTEK), com hidrogénio como gás transportador, a uma taxa de fluxo de 30 ml/min. A temperatura do forno foi inicialmente mantida a 40ºC por 1 min e depois aumentada até 220ºC, numa taxa de 2ºC/min, a qual foi mantida durante 10 min. O detector de ionização por chama foi também mantido a 220ºC.
Análise Estatística
As áreas relativas dos elementos presentes na fracção volátil da análise por CG foram usadas para a análise de dados. O processamento destes dados foi realizado com o programa PASW Statistics 18 (IBM, USA). Aos dados resultantes da cromatografia gasosa, foi realizada uma análise de componentes principais (PCA) que é uma análise estatística multivariada que permite a redução dos dados ao explicar a variância existente entre os componentes principais, ou seja, combinações lineares das variáveis. Deste modo, este tipo de análise permitiu verificar se amostras correspondentes a determinado tratamento produzem uma resposta suficientemente semelhante para se agruparem e se se cria um padrão que as diferencie dos outros grupos (SUMNER et al., 2003).
Todas as outras análises e gráficos, respeitantes aos dados do conteúdo de água e extracção de clorofilas, foram obtidos com Microsoft Excel e GraphPad Software (La Jolla, USA). A normalidade dos dados foi confirmada e o teste paramétrico t-student foi escolhido para comparar os grupos em estudo.
Resultados e discussão
Determinação do conteúdo de água
A infecção causada pelo nemátode bloqueia a condução da água e reduz a taxa de transpiração (TAN et al., 2005). Três horas após a invasão com o nemátode, o conteúdo de água do caule, quer em P. pinaster que em P. pinea diminuiu em comparação com as árvores controlo (Figura 1).
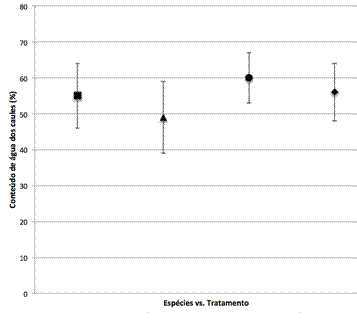
Figura 1 – Conteúdo de água do caule em P. pinaster inoculado com água (n) ou B. xylophilus estirpe HF (p) e em P. pinea inoculado com água (l) ou B. xylophilus estirpe HF (u). Cada valor representa a média ±desvio padrão.
Os resultados demonstraram uma diminuição do conteúdo da água de 10,9% após inoculação com o nemátode em P. pinaster (p<0.05) e de 6,7% em P. pinea (p>0.05), sendo que apenas as diferenças entre P. pinaster inoculado com nemátode e controlo é que foram estatisticamente significativas. O P. pinea como hospedeiro da DMP é ainda muito pouco estudado assim como o porquê desta espécie parecer ter uma menor susceptibilidade ao nemátode (MOTA e VIEIRA, 2008). Mas estes resultados expressam uma clara diferença na resposta à infecção com nemátode entre P. pinaster e P. pinea (p<0.05) o que pode indicar que, no que se refere ao conteúdo de água nos caules, o pinheiro manso é menos afectado do que o pinheiro bravo, o hospedeiro principal da doença em Portugal. Isto pode sugerir, como denotado por FUTAI (2003), que a resistência de certas espécies, como o P. taeda, pode ser parcialmente explicada por certos repelentes hidrofílicos que fazem parte da madeira deste tipo de árvores, que levam à agregação das espécies de Bursaphelenchus às substâncias hidrofóbicas, enquanto a sua invasão nos tecidos hospedeiros são controlados por estas referidas substâncias hidrofílicas.
O desenvolvimento dos sintomas a temperaturas elevadas depende do grau de avanço da cavitação dos traqueídeos causada pela movimentação do nemátode (RUTHERFORD et al., 1992). Os estudos de TAN et al. (2005) em pinheiro negro japonês (P. thunbergii), com 3-4 anos de idade, foram conduzidos entre 5 e 70 dias após infecção (dai) e registaram uma diminuição percentual de 7,2% e 36,1%, a 10 e a 70 dai, respectivamente. Estudos prévios em P. sylvestris e P. strobus com 20-30 anos (BOLLA et al., 1986) durante 3 meses, demonstraram que estas alterações no conteúdo de água podem ser provocadas pelo bloqueio no sistema de transporte de água das plantas, resultante da infecção com o nemátode.
Considerando que este é um estudo nas primeiras horas após infecção, a diminuição do conteúdo de água foi muito aguda e indica que há uma resposta forte e imediata à infecção. Métodos para detectar a DMP no seu estádio inicial são urgentes para evitar a disseminação entre árvores. Como demonstrado na Figura 1, 3 horas após infecção (hai) os conteúdos de água em P. pinaster diminuíram e pode revelar este sintoma como um possível indicador precoce para uma árvore infectada.
Extracção e quantificação das clorofilas totais
A clorose em agulhas é um sintoma distintivo da fase avançada da DMP (FUKUDA, 1997). Na Figura 2 encontram-se os resultados da concentração de clorofilas em P. pinaster e P. pinea, inoculados com nemátode e controlos.
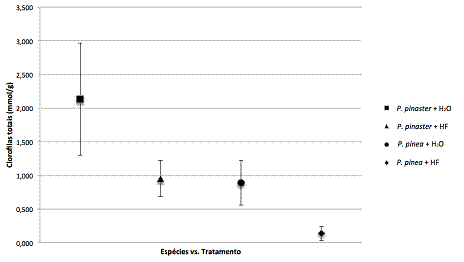
Figura 2 – Concentração das clorofilas totais de P. pinaster inoculado com água (n) ou B. xylophilus estirpe HF (p) e de P. pinea inoculado com água (l) ou B. xylophilus estirpe HF (u). Cada valor representa a média ± desvio padrão
Existe uma variação natural entre a pigmentação das agulhas de P. pinea e P. pinaster, sendo mais escura em P. pinaster. Este facto está relacionado com a maior concentração de clorofilas totais que foram extraídas desta espécie, quando comparadas com P. pinea (p<0.0001).
Mais ainda, após infecção, a concentração de clorofilas diminuiu em ambas as espécies, mais acentuadamente em P. pinaster. Visualmente, nenhuma alteração foi identificada na cor das agulhas dos pinheiros após inoculação, mais especificamente, nenhum indício de amarelecimento das agulhas foi detectado. No entanto, como demontrado na Figura 2, após extracção, houve uma diferença significativa nos níveis de clorofila entre pinheiros inoculados com nemátode e os controlos, o que indica que logo 3 hai os níveis de clorofila começam rapidamente a diminuir, de forma mais abrupta em P. pinaster. Estas diferenças entre tratamentos na concentração de clorofilas totais foram estatisticamente suportadas (p<0.0001) e demonstram que os valores obtidos não são um falso positivo, provocado apenas pelas feridas realizadas para inocular as soluções. Também as respostas entre espécies foram muito diferentes (p<0.0001).
Assim, estes resultados indicam que a diminuição da concentração de clorofila nas agulhas dá-se num estádio precoce da doença. Deste modo, verificou-se que os níveis de clorofila diminuem rapidamente nas primeiras horas pós infecção, o que influencia os processos fotossintéticos e conduz à morte da árvore.
Análise de compostos voláteis
Os voláteis das plantas podem ser emitidos constitutivamente ou podem ser induzidos sob determinadas condições ambientais e têm um papel importante na mediação das interacções entre plantas e insectos herbívoros (MATEUS et al., 2010).
Em estudos prévios (dados não publicados) sobre a expressão genética em P. pinaster e P. pinea 3hai, foram isolados genes relacionados com a produção de metabolitos secundários. Isto levou à utilização de cromatografia gasosa para comparar os padrões dos voláteis emitidos por estas árvores inoculadas com água ou nemátode, no mesmo tempo de amostragem acima referido. No sentido de perceber o padrão das intercorrelações entre as variáveis em estudo (agrupamento de amostras por espécie de pinheiro e/ou por tipo de tratamento), foi conduzida uma análise estatística exploratória, usando a análise de componentes principais (PCA) e o modelo resultou em dois factores principais significativos, que explicaram 44,9% da variância nos dados, tal como representado na Figura 3.
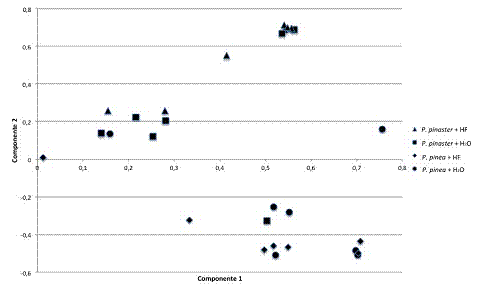
Figura 3 – Análise dos padrões de voláteis de P. pinaster inoculado com água (n) ou B. xylophilus estirpe HF (p) e de P. pinea inoculado com água (l) ou B. xylophilus estirpe HF (u). Um total de 44,9% da variância dos dados foi explicada por dois componentes principais significativos (24,6% da variância na matriz X e 20,3% da variância na variável Y)
Dois grupos foram diferenciados, sendo que a maioria das amostras de P. pinaster foram separadas das de P. pinea pelo factor discriminante com 20,3% de peso na explicação da variância. O primeiro factor discriminante, com 24,6% de peso, não resultou em nenhuma tendência pronunciada entre tratamentos (controlo versus inoculação com nemátode).
SANTOS et al. (2006) realizaram análises de GC-MS de forma a verificar se seria possível distinguir amostras de floema de P. halepensis, P. sylvestris, P. pinaster e P. pinea provenientes da Serra da Lousã, de Alcácer do Sal e da Serra da Ota, e concluíram, tal como no presente trabalho, que os padrões de cada espécie são diferenciáveis e distintos. Adicionalmente, estudos de GCxGC realizados em agulhas de 11 espécies diferentes de pinheiro adulto, localizados em Portugal Central, resultaram numa discriminação das espécies e caracterização química bem sucedidas (MATEUS et al., 2010). MUMM et al. (2003; 2004) analisaram também os voláteis produzidos após deposição de ovos ou feridas mecânicas emitidos quer pelas agulhas de P. sylvestris com 35 a 45 anos, quer pelos ramos de P. nigra de 14 anos de idade, com GC-MS. Perceberam que estes danos afectam significativamente a composição quantitativa da mistura dos voláteis.
Estes resultados indicam que, apesar de ser possível fazer uma distinção clara entre espécies, a produção de voláteis de novo em resposta à infecção com NMP, 3 hai, não produz um padrão diferenciável numa simples corrida de GC, apesar da sua acção como agentes primários na defesa contra patogéneos.
Conclusões
Apesar das modificações fisiológicas analisadas serem normalmente associadas aos estádios tardios da infecção, foi detectada uma tendência para a redução no conteúdo da água e na concentração de clorofilas 3 h após infecção. Este resultado é bastante revelador, dado que a infecção provocou danos quase de imediato na árvore. Também se pode verificar que as respostas foram distintas entre P. pinea e P. pinaster, o que é suportado pela diferença de susceptibilidade entre espécies à doença, já demonstrado por FUTAI (2003) e que pode estar relacionado com as relações filogenéticas dentro do seu género.
Referências bibliográficas
ABADÍA, J., MONGE, E., MONTAÑES, L., HERAS, L., 1984. Extraction of iron from plant leaves by Fe(II) chelators. Journal of Plant Nutrition 7(1-5): 777-784. [ Links ]
AGUIAR, A., ALMEIDA, M.H., BORRALHO, N., 2003. Genetic Control of Growth, Wood Density and Stem Characteristics of Pinus pinaster in Portugal. Silva Lusitana 11(2): 131-139. [ Links ]
BAERMANN, G., 1917. Eine einfache Methode zur Auffindung von Ankylostomum (nematoden) Larven in Erdproben. Geneesk, Tijdschr, Ned-Indie 57: 131-137. [ Links ]
BALDO, A., NORELLI, J.L., FARRELL Jr. R., BASSET, C.L., ALDWINCKLE, H.S., MALNOY, M., 2010. Identification of genes differentially expressed during interaction of resistant and susceptible apple cultivars (Malus x domestica) with Erwinia amylovora. BMC Plant Biology 10(1). doi:10.1186/1471-2229-10-1 [ Links ]
BOLLA, R.I., WINTER, R.E.K., FITZSIMMONS, K., LINIT, M.J., 1986. Pathotypes of the Pinewood Nematode Bursaphelenchus xylophilus. Journal of Nematology 18(2): 230-238. [ Links ]
CORREIA, I., ALMEIDA, H., AGUIAR, A., 2004. Variabilidade do Crescimento e da Forma de Proveniências de Pinus pinaster Aiton aos 8 Anos, na Mata Nacional do Escaroup.Silva Lusitana 12(2): 151-182. [ Links ]
DWINELL, L.D., 1997. The Pinewood Nematode: Regulation and Mitigation. Annu Rev Phytopathol 35: 153-166. [ Links ]
FUKUDA, K., 1997. Physiological Process of the Symptom Development and Resistance Mechanism in Pine Wilt Disease. J For Res 2: 171-181. [ Links ]
FUTAI, K., 2003. Pine Wilt Disease: Various biological relationships and resulting events. Proceedings: International Union of Forest Research Organizations Kanazawa 2003 "Forest Insect Population Dynamics and Host Influences". [ Links ]
FUTAI, K., FURUNO, T., 1979. The variety of resistances among pine species to pine wood nematode, Bursaphelenchus lignicolus. Bull Kyoto Uni For 51: 23-36. [ Links ]
GLEASON, M., LINIT, M., ZRIBA, N., DONALD, P., TISSERAT, N., GIESLER, L., 2000. Pine Wilt: A fatal disease of exotic pines in the Midwest. Sustainable Urban Landscape 9. [ Links ]
KIM, J., SEO, S-M., LEE, S-G., SHIN, S-C., PARK, I-K., 2008. Nematicidal activity of plant essential oils and components from Coriander (Coriandrum sativum), Oriental Sweetgun (Liquidambar orientalis), and Valerian (Valeriana wallichii) essential oils against Pine Wood Nematode (Bursaphelenchus xylophilus). J Agric Food Chem 56: 7316-7320. [ Links ]
KOUTROUMPA, F.A., SALLE, A., LIEUTIER, F., ROUX-MORABITO, G., 2009. Feeding and oviposition preferences of Monochamus galloprovincialis on its main hosts Pinus sylvestris and Pinus pinaster. Entomologia Hellenica 18: 35-46. [ Links ]
MATEUS, E., BARATA, R.C., ZROSTLÍKOVÁ, J., GOMES DA SILVA, M.D.R., PAIVA, M.R., 2010. Characterization of the volatile fraction emitted by Pinus spp. by one- and two-dimensional chromatographic techniques with mass spectrometric detection. Journal of Chromatography A 1217: 1845-1855. [ Links ]
MOTA, M.M., BRAASCH, H., BRAVO, M.A., PENAS, A.C., BURGERMEISTER, W., METGE, K., SOUSA, E., 1999. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1(7-8): 727-734. [ Links ]
MOTA, M.M., VIEIRA, P.R. (Eds.) 2008. Pine Wilt Disease: A worldwide threat to forest ecosystems. Springer, s.l.. 405 p. [ Links ]
MUMM, R., SCHRANK, K., WEGENER, R., SCHULZ, S., HILKER, M., 2003. Chemical analysis of volatiles emitted by Pinus sylvestris after induction by insect oviposition. Journal of Chemical Ecology 29(5): 1235-1252 [ Links ]
MUMM, R., TIEMANN, T., SCHULZ, S., HILKER, M., 2004. Analysis of volatiles from black pine (Pinus nigra): significance of wounding and egg deposition by herbivorous sawfly. Phytochemistry 65: 3221-3230. [ Links ]
MYERS, R.F., 1988. Pathogenesis in Pine Wilt caused by Pinewood Nematode, Bursaphelenchus xylophilus. Journal of Nematology 20(2): 236-244. [ Links ]
NAVES, P., SOUSA, E., QUARTAU, J.A., 2007. Winter dormancy of the pine sawyer Monochamus galloprovincialis (Col., Cerambycidae) in Portugal. J Appl Entomol 131: 669-673. [ Links ]
NICKLE, W.R., GOLDEN, A.M., MAMIYA, Y., WERGIN, W.P., 1981. On the taxonomy and morphology of the Pine Wood Nematode, Bursaphelenchus xylophilus (Steiner & Buhrer 1934) Nickle 1970. Journal of Nematology 13(3): 385-392. [ Links ]
PROENÇA DA CUNHA, A., 2005. Farmacognosia e Fitoquímica. Fundação Calouste Gulbenkian, Lisboa. 671 p. [ Links ]
RUTHERFORD, T.A., RIGA, E., WEBSTER, J.M., 1992. Temperature-mediated behavioural relationships in Bursaphelenchus xylophilus, B. mucrunatus, and their hybrids. Journal of Nematology 24(1): 40-44. [ Links ]
SANTOS, A.M., VASCONCELOS, T., MATEUS E., FARRALL, M.H., GOMES DA SILVA, M.D.R., PAIVA, M.R., BRANCO, M., 2006. Characterization of the volatile fraction emitted by phloems of four pine species by solid-phase microextraction and gas chromatography-mass spectrometry. Journal of Chromatography A 1105: 191-198. [ Links ]
SIKORA, E.J., MALEK, R.B., 1991. Influence of temperature on development of Pine Wilt in Scots Pine. Journal of Nematology 23(2): 188-193. [ Links ]
SUMNER, L.W., MENDES, P., DIXON, R.A., 2003. Plant metabolomics: large-scale phytochemistry in the functional genomics era. Phytochemistry 62: 817-836. [ Links ]
TAN, J-J., YE, J-R., WU, X-Q., ZHU, Y-F., LI, Y., 2005. A study on disease development and early diagnosis of pine wood nematode, Bursaphenchus xylophilus, infection of Japanese black pine. Nematology 7(4): 481-485. [ Links ]
ZHAO, B.G., FUTAI, K., SUTHERLAND, J.R., TAKEUCHI, Y. (Eds.), 2008. Pine Wilt Disease. Springer, s.l.. 459 p. [ Links ]
ZHAO, L., WEI, W., LIU, X., KANG, L., SUN, J., 2007. A novel rapid sampling method for pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Parasita-phelenchidae). Can. J. For. Res. 37: 1867-1872. [ Links ]
Agradecimentos
Os autores gostariam de agradecer à Autoridade Florestal Nacional e ao Instituto de Financiamento da Agricultura e Pescas, I.P., pelo financiamento do projecto "Estratégia biotecnológica para combater a doença da madeira do pinheiro", Dr. Manuel Mota e Dr. Pedro Barbosa por terem cedido a estirpe de nemátode HF e à Doutora Marta R. M. Lima pelo apoio na técnica de cromatografia gasosa.
Entregue para publicação em Março de 2011
Aceite para publicação em Maio de 2011
