










Serviços Personalizados
Journal
Artigo
 Francês (pdf)
Francês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkSilva Lusitana
versão impressa ISSN 0870-6352
Silva Lus. v.18 n.1 Lisboa jun. 2010
Apoidea et leur Diversité au Nord d'Algérie
Leila Bendifallah*, Kamel Louadi** et Salah Eddine Doumandji**
*PhD, Départment de Biologie. Faculté des Sciences. Université M'hamed Bougara. Boumerdes, ALGÉRIE, E-mail: bendif_L@yahoo.fr
**Professeur, Laboratoire de Biosystématique et Ecologie des Arthropodes. Faculté des Sciences de la Nature et de la Vie. Université Mentouri. Constantine, ALGÉRIE / Laboratoire de Zoophytiatrie. Département de Zoologie Agricole et Forestière. École Nationale Supérieure Agronomique d'El Harrach – Alger, ALGÉRIE
Résumé
Les abeilles sont les sentinelles de l'environnement. Elles contribuent à la biodiversité des plantes et à la sécurité alimentaire. La présente étude concerne la faunistique et la biodiversité des abeilles sauvages dans l'écosystème naturel. Les investigations sont menées au niveau de quatre localités: la Mitidja centrale représentée par El Harrach, Boumerdes, Bouira et Blida durant les périodes allant de 2004 à 2008. Un total de 4300 spécimens a été pris en compte, 120 espèces d'abeilles sauvages sont présentes dans les régions d'études. La faune d'abeilles sauvages recensée est répartie entre 4 familles et 20 genres. Une importante diversité est notée pour la famille des Halictidae. Quatre espèces et 3 sous-espèces nouveaux registres sont enregistrées pour les régions d'étude. Le maximum de taxa est observé au mois d'avril coïncidant avec la floraison de la majorité des plantes. La densité et la diversité des abeilles sont différentes d'une station à une autre selon les facteurs climatiques et la disponibilité des ressources végétales. La station d'El Harrach (Mitidja) est un terrain expérimental destiné au maraîchage alors que la station de Blida est destinée à la plantation de l'avoine, celle de Boumerdes est laissée en jachère et la station de Bouira située sur une montagne n'est destinée à aucune plantation mais elle est entourée d'une oliveraie.
Mots clés: Abeilles sauvages; diversité; écosystème naturel; Nord d'Algérie.
A Diversidade de Apoidea no Norte da Argélia
Sumário
As abelhas são as sentinelas do ambiente. Contribuem para a biodiversidade das plantas e para a segurança alimentar. O presente estudo refere-se à faunística e biodiversidade das abelhas selvagens num ecossistema natural. A investigação desenvolve-se em quatro localidades: na Mitidja Central, representada por El Harrach, Boumerdes, Bouira e Blida, em períodos compreendidos entre 2004 e 2008. Foram considerados 4300 exemplares, existindo 120 espécies de abelhas selvagens nas regiões de estudo. A fauna de abelhas selvagens recenseadas distribui-se por 4 famílias e 20 géneros. À família Halictidae corresponde uma diversidade considerável. Nas regiões estudadas registaram-se 4 espécies e 3 sub-espécies novas. O número máximo de taxa observa-se no mês de Abril, coincidindo com a floração da maioria das plantas. A densidade e diversidade das abelhas varia de uma estação para outra, segundo os factores climáticos e a disponibilidade de recursos vegetais. A estação de El Harrach (Mitidja) é um terreno experimental destinado a horticultura, na estação de Blida planta-se aveia, Boumerdes está em poisio e a estação de Bouira, localizada numa montanha, não se destina a plantação, mas está rodeada de olival.
Palavras-chave: Abelhas selvagens; diversidade; ecossistema natural; Norte da Argélia
Apoidea and their Diversity in the North of Algeria
Abstract
Bees are the sentinels of the environment. They contribute to plant biodiversity and food security. This study concerns the wildlife and biodiversity of wild bees in a natural ecosystem. Investigations are conducted at four locations: the Central Mitidja represented by El Harrach, Boumerdes, Blida and Bouira during the periods from 2004 to 2008. A total of 4300 specimens was taken into account, with 120 species of wild bees present in the study areas. The fauna of wild bees recorded is divided between 4 families and 20 genera. A significant diversity is noted for the Halictidae family. Four species and three subspecies are new records for the study areas. The maximum number of taxa is observed in April coinciding with the flowering of most plants. The density and diversity of bees are different from one station to another depending on climatic factors and food availability. The station of El Harrach (Mitidja) is an experimental field for legume crops, while the station of Blida is intended for planting of oat, the Boumerdes station is for grazing and the station of Bouira, located on a mountain, is not intended for planting but is surrounded by an olive grove.
Key words: Wild bees; diversity; natural ecosystem; North of Algeria
Introduction
Les abeilles domestiques et sauvages revêtent un grand intérêt au niveau des écosystèmes naturels et de l'agro-cénose. En effet, beaucoup de travaux montrent que les abeilles sont les meilleurs agents pollinisateurs (McGREGOR, 1976). Probablement, leur activité la plus importante, en termes d'avantages pour l'homme, est leur pollinisation de la végétation naturelle ( MICHENER , 2007). La pollinisation est l'un des mécanismes les plus importants dans le maintien et la promotion de la diversité biologique et, en général, de la vie sur terre. En outre, un tiers des cultures nécessite une pollinisation pour améliorer la qualité des graines et des fruits et la grande majorité d'entre elles sont pollinisées par de nombreuses abeilles estimées à 25.000 espèces ( DIAS et al., 1999). La faunistique et la diversité des abeilles sauvages ont été largement étudiées ( JAVOREK et al., 2002; DUPONT et SKOV, 2004; MICHEZ et al., 2004c; KUHLMAN, 2005; POTTS et al., 2005; BODIN et al., 2006; STEFFAN-DEWENTER et al., 2006; BIESMEIJER et al., 2006; KLEIN et al., 2007; PATINY et MICHEZ, 2007; MÜLLER et KUHLMANN , 2008). À l'échelle Ouest-paléarctique, on observe la plus grande diversité en apoïdes dans la région méditerranéenne (MICHENER, 1979; RASMONT et al., 1995). Néanmoins, trop peu d'inventaires faunistiques et floristiques sont réalisés en région du Maghreb (Afrique du Nord), pourtant ils constituent l'élément essentiel pour la détermination de la biodiversité d'un écosystème et la gestion de celui-ci. Selon RASMONT et al. (1995), cette zone présente probablement une diversité très élevée similaire ou plus grande que celle de la Californie. En Algérie , les études réalisées jusqu'à présent sur les Apoidea sont celles de SAUNDERS (1901, 1908), ALFKEN (1914), SCHULTHESS (1924), ROTH (1923, 1924 et 1930) et traitent seulement une petite partie de l'Algérie. Les travaux les plus récents sont ceux effectués dans la région Est de l'Algérie ( LOUADI et DOUMANDJI , 1998a, 1998b; LOUADI , 1999a, 1999b; BENACHOUR 2007; LOUADI et al., 2007a, 2007b, 2008) et à Tizi Ouzou ( AOUAR- SADLI et al., 2008).
L'Algérie est un pays soumis à l'influence conjuguée de la mer, du relief et de l'altitude. Le climat est de type méditerranéen tempéré. Il est caractérisé par une longue période de sécheresse estivale variant de 3 à 4 mois sur le littoral, de 5 à 6 mois au niveau des Hautes Plaines, et supérieure à 6 mois au niveau de l'Atlas Saharien (SELTZER, 1946 ; STEWART, 1969). Avec ses six régions écologiques, l'Algérie possède une grande diversité floristique. Les températures annuelles et les précipitations diffèrent d'une région à une autre. En effet, les températures annuelles moyennes sont comprises entre 11 et 20°C. et les pluviométries annuelles sont comprises entre 500 et 1300 mm. La présente étude veut apporter un nouvel éclairage sur la connaissance des Apoidea sauvages à travers quelques localités du Nord d'Algérie. Les aspects essentiels traités sont (1) l'établissement de la liste des espèces d'apoïdes au Nord d'Algérie, (2) la biogéographie de la faune d'abeilles et sa diversité à travers les différentes régions d'étude (3) et valider ou non l'hypothèse concernant la faible diversité des apoïdes solitaires dans les régions montagnardes par rapport à celles situées en basse altitude.
Matériel et méthodes
La biogéographie et la répartition des espèces d'abeilles sont traitées dans un transect Est - Ouest des régions du centre d'Algérie: Bouira (Ahl El Ksar), Mitidja orientale (El Harrach), Boumerdes et Blida (Figure 1). Le travail est réalisé dans le milieu naturel sur des plantes spontanées durant les périodes hivernale, printanière et estivale des années 2004 à 2007.
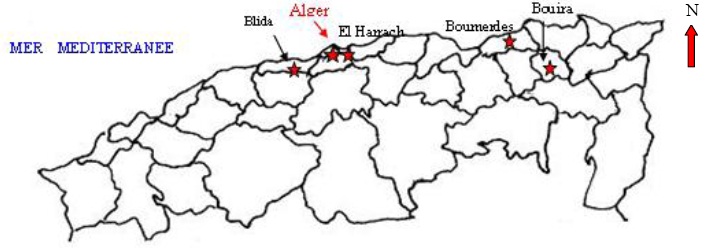
Figure 1 – Localisation des stations d'étude
Les données bioclimatiques dans les régions étudiées sont différentes (Tableau 1).
Tableau 1 - Caractéristiques des quatre stations d'étude. T°moy. ann.: température moyenne annuelle, Précip. ann. (mm): précipitations moyennes annuelles

Composition
Pour identifier la faune des abeilles présentes dans les milieux d'étude, plusieurs méthodes sont employées afin de récolter le maximum d'espèces. Aussi, certaines techniques sont spécifiques à quelques familles d'abeilles telles que la chasse à vue. Celle–ci consiste à capturer les grosses abeilles comme les Xylocopinae, les Anthophoridae et les Bombinae à l'aide d'un filet entomologique (SONET et JACOB-REMACLE, 1987). L'échantillonnage se réalise pendant une demi journée de 8 heures à 12 heures.
Une autre technique non moins importante que la précédente, est utilisée pour l'échantillonnage et la capture des abeilles: la chasse à l'aide des sachets ou tubes en matière plastique. Cette technique seffectue par approche directe avec des tubes en matière plastique de 5 cm. de hauteur et 3 cm de diamètre contenant un papier filtre imbibé de quelques gouttes d'éther-acétique (GUIGLIA, 1972, LOUADI et DOUMANDJI, 1998b). Cette méthode est très pratique, elle permet de capturer les espèces les plus rapides et beaucoup d'espèces de petite taille.
Une autre méthode est celle de l'aspirateur à bouche. Ce dernier est conçu avec un bocal et deux tuyaux flexibles. La dernière méthode de capture est celle des pièges à eau colorée.
Comptage des Apoidea
Elle est réalisée pendant la période printanière. Celle–ci coïncide avec la floraison d'un maximum de plantes et le vol de la majorité des abeilles. A partir du 29 mars jusqu'au 28 juin, 8 observations sont faites par journée, de 7 h. à 15 h. (Gmt + 1). L'observateur compte les abeilles à chaque heure de la journée. Les comptages s'effectuent deux fois par semaine. Cette méthode de comptage est celle du transect végétal adaptée aux plantes herbacées ( TASEÏ , 1976; PARKER, 1981). Il s'agit de 5 transects, avec 1m de large, correspondant à 2 rectangles contigus ayant 380m de contour:
2 × 100m + 3 × 60m.
Les données et les résultats obtenus à travers toute étude doivent être exploités en adoptant un certain nombre de méthodes. Celles-ci sont spécifiques à chaque méthode d'échantillonnage et aux objectifs visés. SOUTHWOOD (1978) propose pour l'étude des communautés animales, notamment des insectes, d'effectuer des analyses de distribution d'abondance et des indices écologiques tels que la diversité. Nos résultats sont exploités dans cet axe.
Analyse des données
La diversité spécifique peut être quantifiée par différents indices mathématiques. Dans notre cas, nous avons choisi d'en utiliser l'indice de Shannon-Weaver et l'espérance de Hurlbert.
- Indice de Shannon-Weaver
Il estime la diversité spécifique en quantité d'information. Il s'exprime en bit.
H' = — ∑ piLog2 Pi, où
H' est l'indice de diversité exprimé en unité bits.
Pi = ni/N
ni est le nombre de spécimens de l'espèce i.
N est le nombre total des spécimens de la station
Log2 est le logarithme à base 2
- Espérance de Hulbert
Il représente le nombre d'espèces espérées dans un tirage aléatoire de 100 spécimens et ce pour le site d'échantillonnage. Il s'exprime en espèce/100 spécimens. L'Es évalue la diversité d'un site. La formule simplifiée par RASMONT et al. (1990).
![]()
ni: nombre de spécimens de l'espèce i
N: nombre total de spécimens de la station
- Equirépartition
Cet indice permet de savoir comment se fait la répartition des effectifs entre les diverses espèces présentes. Selon RAMADE (1984), les valeurs de E varient entre 0 et 1.
Gestion des données
Les données sont gérées à l'aide du logiciel Data Fauna Flora (DFF version 2.0) ( BARBIER et al., 2002). Pour comparer l'abondance des taxa entre différentes stations, un test statistique de Kruskal-Wallis, a été utilisé.
Pour comparer l'abondance entre les différents taxa et entre les sites, une analyse de la variance par le modèle linéaire global ( G.L.M .) a été employée. Cette analyse est faite par le programme SYSTAT vers. 12, SPSS 2009.
Résultats et discussion
Composition et diversité spécifique des Apoidea
L'étude de la biodiversité des abeilles sauvages solitaires et sociales a mis en évidence 120 espèces recensées au Nord d'Algérie au niveau de quatre régions détude sur un total de 4300 spécimens observés répartis dans 20 genres (Tableau 2). Cette étude a permis de noter 4 nouvelles espèces et 3 nouvelles sous-espèces pour la faune des Apoides du Nord d'Algérie qui n'ont pas été citées par ALFKEN (1914). Il s'agit de Bombus (Megabombus) ruderatus Scopoli, 1763 sous-espèce siculus Friese, 1882; Anthophora atriceps Perez, 1879; Xylocopa (Copoxyla) iris Christ, 1791 sous-espèce cupripennis Smith, 1874, Lasioglossum (Lasioglossum) discum Smith, 1853 sous-espèce aegyptiellum et Lasioglossum (Lasioglossum) transitorium. Seules quelques espèces abondent (Andrena flavipes, Andrena florentina, Andrena bimaculata, Halictus scabiosae, Halictus sp1., Halictus rufipes, Lasioglossum zonolum ) et représentent 32% de l'effectif total. 23 taxa sont représentés par un seul spécimen (singleton).
Tableau 2 – Abondance des spécimens par genre et par famille d'Apoidea dans les quatre stations d'étude. Ni = nombre de spécimens
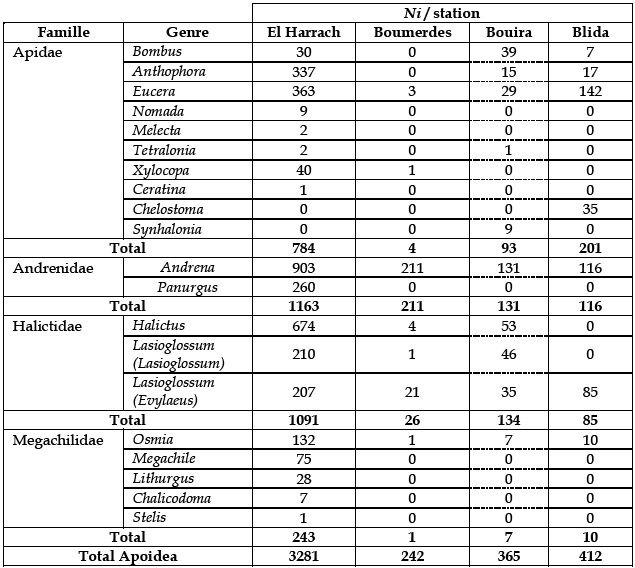
Les familles d'abeilles sauvages sont représentées par 30% des espèces observées appartenant à la famille des Halictidae. Cette famille apparaît comme la famille la plus diversifiée au Nord d'Algérie (Figure 2). Cependant, la famille des Andrenidae est la famille la plus abondante représentant à elle-seule 36,83% des spécimens observés (Figure 3).
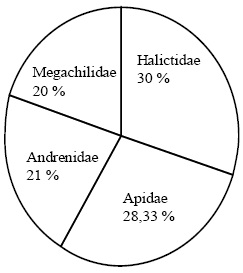
Figure 2 – Diversité des familles d'abeilles sauvages dans les régions d'étude
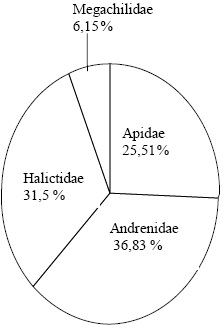
Figure 3 – Abondance des familles d'abeilles sauvages dans les régions d'étude
La famille des Megachilidae est peu diversifiée et peu abondante.
Les familles d'abeilles notées sont les mêmes que celles signalées par Louadi et DOUMANDJI (1998a et b) dans la région de Constantine. Néanmoins les Colletidae et les Mellitidae sont absentes aussi bien de la région du Nord d'Algérie et du Constantinois, bien qu'elles soient recensées en 1914 par Alfken dans la région algéroise au cours de l'automne. Par ailleurs, l'inventaire des abeilles établi au printemps par SONET et JACOB - REMACLE (1987) en Tunisie, pays limitrophe à l'Algérie, n'a pas révélé non plus la présence de ces deux familles.
Généralement, la famille la plus diversifiée en région Ouest-paléarctique est celle des Halictidae ((GONZALEZ et al., 1999 (Espagne), OERTLI et al., 2005 (Suisse), LACHAUD et MAHE, 2008 (France), STALLEGGER et LIVORY, 2008 (France), BARONE et al., 1999 (Belgique) et MONSEVIEIUS, 2004 (Lithuanie)). En ce qui concerne le nombre des spécimens, les familles les plus abondantes sont les Halictidae et les Andrenidae. La présente étude présente des résultats similaires, une abondance notable est enregistrée pour les familles des Andrenidae et des Halictidae (Tableau 2).
Distribution d'abondance des espèces d'Apoidea dans les quatre stations d'étude
La représentation graphique de l'abondance-dominance des taxa (Figure 4) semble indiquer une diversité notable au Nord d'Algérie. Comme précédemment, on observe la très nette dominance de trois taxa: Andrena flavipes, Andrena florentina et Halictus scabiosae par rapport aux autres taxa.
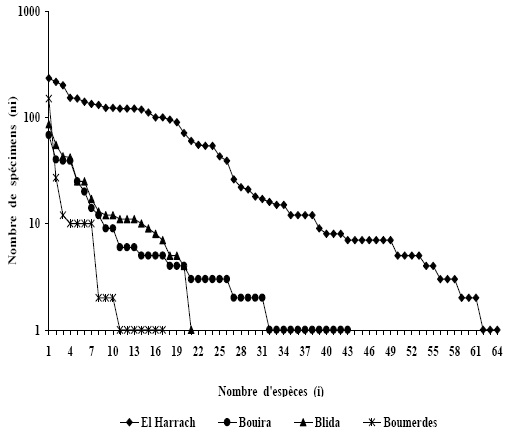
Figure 4 – Représentation graphique de la distribution d'abondance des espèces d'Apoidea, i en abscisse et ni en ordonnées pour les quatre stations d'étude
La station d'El Harrach est plus diversifiée en nombre de taxa et d'effectifs par rapport aux stations de Boumerdes, Blida et Bouira. Ceci peut résulter d'un sur-échantillonnage. De plus, les différences de diversités spécifiques constatées peuvent s'expliquer par des périodes d'échantillonnage différentes. L'inventaire faunistique réalisé à El Harrach a été mené sur des périodes d'observations longues, 16 mois, avec 3281 spécimens. La faible proportion de singleton dans cette étude (3 taxa) suggère que ce site a été sur-échantillonné ou que les autres sites ont été sous-échantillonnés. À l'inverse, on observe d'importantes proportions en singleton en régions de Bouira (11 taxa) et Blida (7 taxa). Des études similaires réalisées par LACHAUD et MAHE (2008) et de GONZALEZ et al. (1999) montrent un nombre d'espèces représentées par un spécimen relativement important. Ce qui laisse présager que ces sites présentent soit une certaine originalité faunistique avec la présence de nombreuses espèces rares, soit un sous-échantillonnage.
Les Apidae sauvages Bombus terrestris et Bombus ruderatus siculus sont en nombre important sur le site de Bouira comparés aux autres sites, ceci s'explique par le fait que cette station est située en haute altitude et que les bourdons pourraient être particulièrement bien adaptés aux milieux montagnards (ISERBYT et al., 2008). En outre, les ressources trouvées sur ce site telles que Rosmarinus officinalis L., Cistus ablidus L., Glladiolis segetum L. Leontodon hispidus L. et Galactites tomentosa L.sont appréciées par les bourdons. Quelques faits semblent appuyer cette hypothèse dont notamment l'endothermie qui est plus élaborée chez les langues longues que chez les langues courtes ( ALFORD, 1975; HENRICH, 1979). LOUADI DOUMANDJI (1998a) constatent la même observation à Constantine, région de haute altitude de 660 m. Ils notent un effectif de 130 spécimens pour les deux espèces de bourdons.
D'après MICHENER (1979) et PITKÄNEN et TIAINEN (2001), la région méditerranéenne est la plus riche et la plus diversifiée pour les abeilles solitaires. Cette diversité diminue ( PITKÄNEN et TIAINEN, 2001) en fonction de la latitude et par conséquent de l'altitude.
Malgré le faible nombre d'inventaires des abeilles solitaires en général, la diversité des abeilles solitaires semble diminuer en fonction de la latitude contrairement aux abeilles sociales. En effet, un gradient en latitude avec une diminution de la diversité vers le nord est constaté (108 espèces en Loire-Atlantique (LACHAUD et MAHE, 2008), 86 espèces en Belgique ( BARONE et al., 1999) et 74 espèces aux Pays-Bas ( PEETERS et REEMERS, 2001). La plus faible diversité est observée dans la région de Cumbria en Angleterre avec 26 espèces (ARCHER, 2008). Comme l'Algérie se situe au Sud de la Méditerranée, une augmentation de la diversité des abeilles solitaires avec le gradient de la latitude est observée. En effet, un total de 120 taxa est inventorié uniquement dans quatre régions d'étude. Cela suppose qu'une importante diversité d'Apoidea existe à travers toutes les régions d'Algérie. La faible diversité spécifique observée dans notre étude sur le site de Boumerdes (17 taxa) est dû soit au sous-échantillonnage, soit à la proximité du site de la mer. Ce site est un milieu ouvert, localisé à une centaine de mètres de la mer, il est influencé par des vents forts lesquels sont accentués par les vents marins salés qui seront à l'origine de la destruction de la végétation, notamment basse. Par conséquent, l'activité des abeilles sera réduite. A l'inverse, la station d'El Harrach bien qu'elle soit située en basse altitude, présente une diversité spécifique importante (64 taxa). Ceci est dû soit au sur-échantillonnage, soit à la grande richesse floristique du site. En effet, 36 espèces de plantes spontanées sont inventoriées. Ce site est situé dans la station expérimentale de l'École nationale supérieure agronomique, les différents essais sont irrigués, par conséquent le milieu bénéficie d'une humidité remarquable.
Phénologie des familles d'abeilles sauvages
Les Apoidea sont actifs selon les facteurs climatiques et les paramètres interspécifiques et intraspécifiques (Figure 5). Les effectifs des quatre familles varient selon les mois. Les résultats notés dans la station d'El Harrach montrent que la plupart des Apoides sont mieux représentés en mars et en avril. Ceci coïncide à la fois avec la floraison d'un maximum de plantes et avec l'installation de conditions clémentes. En Tunisie, SONET et JACOB-REMACLE (1987) ayant travaillé sur la légumineuse Hedysarum coronarium L. décèlent un grand nombre d'abeilles sauvages en avril et en mai, notamment les Apidae. LOUADI et DOUMANDJI (1998b) notent que la plupart des familles d'Apoidea observées sont très bien représentées au mois d'avril. A Liège (Belgique), où des études similaires sont faites par JACOB-REMACLE (1989), et en France par RASMONT (1995), les Megachilidae, les Andrenidae et les Apidae atteignent leur pic d'abondance au mois d'avril. Les Halictidae sont abondants en juillet et août. Nous observons une certaine contradiction avec nos résultats concernant les Andenidae et les Halitidae. Dans notre présente étude, les Halictidae sont abondants en avril et les Andrenidae en mai. Ainsi, les différences observées entre les régions du Nord d'Algérie et Liège relèvent certainement du climat et de la floraison. En effet, le climat estival en Belgique correspond au climat printanier au Nord d'Algérie.

Figure 5 – Phénologie des familles d'abeilles
Répartition et biodiversité des abeilles sauvages à travers les régions d'étude
La répartition et la biodiversité des espèces d'abeilles sauvages sont traitées selon les indices écologiques (Tableau 3). L'impression de diversité assez importante dégagée par la représentation graphique de la distribution d'abondance des espèces d'Apoidea est confirmée par la valeur importante de l'espérance de Hurlbert dans le site d'El Harrach où on peut espérer 38 espèces dans 100 spécimens pris au hasard. Mais, cette valeur est relativement faible pour les sites de Blida, Bouira et Boumerdes qui présentent respectivement 17, 12 et 10 espèces d'abeilles espérées dans 100 individus. Ce même indice comparé à celui du site de Constantine (17 espèces) (LOUADI et DOUMANDJI, 1998a) renforce l'idée d'une richesse spécifique assez importante notamment sur le site d'El Harrach.
Tableau 3 - Indices de diversité des espèces inventoriées dans les quatre localités Ni: nombre d`exemplaires, H`: Indice de diversité de Shannon-Waever, E:Equitabilité, E sIndice d`Hubert
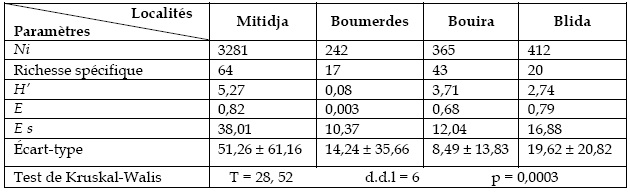
Ces données viennent confirmer la richesse faunistique importante du Nord d'Algérie. Elles appuient l'hypothèse émise par RASMONT et al. (1995b). En effet, d'après ces auteurs, cette diversité serait encore plus grande si on englobe l'ensemble de l'Algérie.
Les tableaux 2 et 3 montrent parfaitement que les différents taxons ainsi que les familles d'abeilles solitaires et sauvages diffèrent par la densité et la diversité selon la station d'étude.
En effet, cette variation du nombre d'espèces est hautement significativement différente.
Le test de Kruskal-Walis donne une approximation par distribution du Ch2 une probabilité égale à 3o avec un T= 28,52 (d.d.l = 6). Ceci signifie que le nombre d'espèces diffère d'une station à une autre et le risque d'erreur est de 3o.
Analyse de la variance entre les sites et les taxa
L'analyse de la variance de l'abondance des abeilles entre les sites démontrée par le test de GLM (Tableau 4, Figure 6) donne F-ratio = 8,700 avec une probabilité inférieur à 10. Ceci confirme une diversité d'abondance hautement significative entre les stations d'étude.
Tableau 4 - Analyse de la variance entre les sites et les taxa par le test de GLM

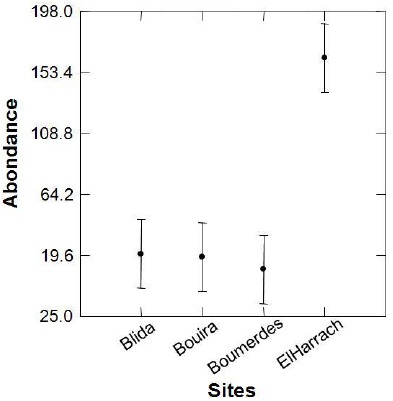
Figure 6 –Différence d'abondance entre les sites d`étude
Concernant les espèces d'Apoidea, l'analyse de la variance de l'abondance des abeilles entre les taxa (Tableau 4, Figure 7) donne F-ratio = 2,129 avec une probabilité inférieur à 15. Ceci confirme une différence significative entre les espèces.

Figure 7 –Différence d'abondance entre les taxa
Les Apoidea se diversifient selon des facteurs climatiques et la disponibilité des plantes préférées caractérisant chaque étage bioclimatique. Des études sur de nombreux groupes taxonomiques ont prouvé que le climat et les facteurs bioclimatiques, comme la température, l'humidité ou l'évapotranspiration, peuvent être corrélés à la richesse en espèces, car ils ont non seulement limité la distribution des espèces, mais aussi la disponibilité des ressources ( ANDREWS et O'BRIEN , 2000; CEBALLOS et BROWN , 1995; GORODKOV 1990, 1992, et PATINY et MICHEZ , 2007). Dans les régions chaudes, la disponibilité de l'eau est un facteur limitant potentiellement puissant pour la distribution des abeilles ( GESS , 1992; MAYER et KUHLMANN , 2004). En effet, le site de Bouira situé dans les hautes plaines est caractérisé par une longue période de sécheresse estivale variant de 5 à 6 mois et des précipitations annuelles moyennes évaluées à 540,6 mm; ces conditions influencent la disponibilité des ressources végétales (8 espèces de plantes) et par conséquent limitent la diversité des abeilles d'où une abondance des espèces relativement moyenne avec un écart-type faible.
La situation des abeilles en général est aujourd'hui très inquiétante. On constate depuis les années 80 un déclin radical de la diversité et des populations (LECLERCQ et al., 1980; WILLIAMS, 1982; RASMONT et MERSCH, 1988; RASMONT et al., 1993, 2005; BIESMIJER et al., 2006). Indispensables à la survie de nos espaces sauvages, à l'agriculture et à l'économie par leur rôle en tant que principaux pollinisateurs, les abeilles constituent un enjeu majeur pour la conservation de la nature.
Ce travail constitue un point de départ de nos recherches futures. Au vue de l'originalité des apoïdes (présente étude), il serait opportun de continuer à établir des inventaires plus exhaustifs à travers toutes les régions d'Algérie, à étudier leur cycle biologique, le comportement alimentaire et leur efficacité pollinisatrice.
Remerciements
Les auteurs expriment leur gratitude aux spécialistes ayant identifié certaines espèces d'abeilles sauvages, Dr. Michael Kuhlmann du Musée d'Histoire Naturelle de Londres, section des hyménoptères, Dr. Stuart P.M. Roberts de l'Université de Reading, Royaume Unie, Professeur Pierre Rasmont, Dr. Michaël Terzo, Dr. Denis Michez et Dr. Stéphanie Iserbyt du Laboratoire de Zoologie de l'Université Mons-Hainaut, Belgique, Dr. Alain Pauly du Département d'Entomologie de l'Institut Royal des Sciences Naturelles de Belgique et Dr. Sébastien Patiny de la Faculté Universitaire des Sciences Agronomiques de Gembloux, Belgique.
Nous remercions sincèrement Dr. Zahr-Eddin Djazouli, Département d'Agronomie, Université Saad Dahleb de Blida, Algérie pour l'analyse statistique des données.
Références
ALFKEN, J.D., 1914. Beitrag zur Kenntnis der Bienenfauna von Algerien. Mém. Soc. Entom. Belgique, T. 22: 185–237.
ALFORD, D.V., 1975. Bumblebees. Davis-Poynter, London, XII + 352 p., 16 pls.
ANDREWS, P., O'BRIEN, E.M., 2000. Climate, vegetation, and predictable gradients in mammal species richness in southern Africa. Journal of Zoology 251: 205-231.
AOUAR-SADLI, M., LOUADI, K., DOUMANDJI, S., 2008. Pollination of the broad bean (Vicia faba L. var. major) (Fabaceae) by wild bees and honey bees (Hymenoptera: Apoidea) and its impact on the seed production in the Tizi-Ouzou area (Algeria). Afr. J. of Agri. Res. 3(4): 266-272.
ARCHER,
BARBIER, Y., RASMONT, P., DUFRENE, M., SIBERT, J.-M., 2002. Data Fauna Flora. Version 2.0.2. Université de Mons-Hainaut, 106 pp. 1CD-ROM.
BARONE, R., RASMONT, P., BARBIER, Y., TERZO, M., 1999. Evaluation faunistique et floristique de la Grande Bruyère de Blaton (Belgique, Hainaut). Université de Mons-Hainaut, 72 pp.
BENACHOUR, K., LOUADI, K., TERZO, M., 2007. Rôle des abeilles sauvages et domestiques (Hymenoptera: Apoidea) dans la pollinisation de la fève (Vicia faba L. var. major) (Fabaceae) en région de Constantine (Algérie). Ann. soc. entomol. Fr. (n.s.) 43(2): 213-219.
BIESMIJER, J.C., ROBERTS, S.P.M., REEMER, M., OHLEMÜLLER, R., EDWARDS, M., PEETERS, T., SCHAFFERS, A.P., POTTS, S.G., KLEUKERS, R., THOMAS, C.D., SETTELE, J., KUNIN, W.E., 2006. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313: 351-354.
BODIN, O., TENGOE, M., NORMAN, A., LUNDBERG, J., ELMQUIST, T., 2006. The value of small size: Loss of forest patches and ecological thresholds in southern Madagascar. Ecological Applications 16: 440-451.
BRAULIO, S.F., DIAS, ANTHONY RAW, VERA, L., IMPERATRI –FONSECA, 1999. International Pollinators Initiative: The São Paulo Declaration on Pollinators. Report on th Recommendations of the Workshop on the Conservation and Sustainable Use of Pollinators in Agriculture with Emphasis on Bees. Brasília: Brazilian Ministry of the Environment (MMA) 1999. 66 pp.
CEBALLOS, G., BROWN, J., 1995. Global Patterns of Mammalian Diversity, Endemism and Endangerment. Conservation Biology 9: 559-568.
DIAS, B.S.F., RAW, A., IMPERATRI – FONSECA, V.L., 1999. International Pollinators Initiative: The São Paulo Declaration on Pollinators. Report on the Recommendations of the Work-shop on the Conservation and Sustainable Use of Pollinators in Agriculture with Emphasis on Bees. Brasília: Brazilian Ministry of the Environment (MMA) 1999. 66 pp.
DUPONT ,Y.L., SKOV, C., 2004. Influence of geographical distribution and floral traits on species richness of bees (Hymenoptera: Apoidea) visiting Echium species (Boraginaceae) of the Canary Islands. International Journal of Plant Sciences 165: 377-386.
GESS, S.K., 1992. Biogeography of the masarine wasps (Hymenoptera: Vespidae: Masarinae) with particular emphasis on the southern African taxa and on the correlations between masarine and forage plant distributions.Journal of Biogeography 19: 491-503.
GONZÀLEZ ,J.A., TORRES, F. GAYUBO, S.F., 1999. Estudio de biodiversidad de abejas (Hymenoptera: Apoidea) en un biotopo arenoso de la Submeseta Norte (España). Zool. Baetica 10: 87-111.
GORODKOV, K.B., 1990. Dynamics of Range: A General Approach I. Entomological Society of America.Entomological review. 69: 110-130. (Traduction from Russian: Entomologicheskoye Obozreniye).
GORODKOV K.B., 1992. Dynamics of Range: General Approach II. Dynamics of Range and Evolution of Taxa (Qualitative or Phyletic Changes of Range). Entomological review 70: 81-99. (Traduction from Russian: Entomologicheskoye Obozreniye).
GUIGLIA, D, 1972. Les guêpes sociales (Hymenoptera, Vespidae) d'Europe occidentale et septentrionale. Ed. Masson et Cie, Parie, 186 pp.
HEINRICH, B., 1979. Bumblebee economics. Harvard University Press, Cambridge, 246 p. [ Links ]
ISERBYT, S., DURIEUX, E.A., RASMONT, P., 2008. The remarkable diversity of bumblebees (Hymenoptera: Apoidea: Bombus) in the Eyne Valley (France, Pyrénées-Orientales). Ann. soc. entomol. Fr 44(2): 211-241 .
JACOB-REMACLE, A., 1999. Abeilles sauvages et pollinisation. Unit. Zool. Génér. Appl., Fac. Sci. Agro. Gembloux pp. 1-40.
JAVOREK, S.K., MACKENZIE, K.E., VANDER KLOET, S.P ., 2002. Comparative pollination effectiveness among bees (Hymenoptera: Apoidea) on lowbush blueberry (Ericaceae: Vaccinum angustifolium). Annals of the Entomological Society of America 95: 345-351.
KUHLMAN. M., 2005. Diversity, Distribution Patterns and Endemism of Southern African Bees (Hymenoptera: Apoidea). In African Biodiversity: Molecules, Organisms, Ecosystems. Eds (Bernard A. Huber, Bradley J. Sinclair and Karl-Heinz Lampe). pp. 167-172.
KLEIN, A.M., VAISSIERE, B.E., CANE, J.H., STEFFAN-DEWENTER, I., CUNNINGHAM, S.A., KREMEN, C., TSCHARNTKE, T., 2007. Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society of London 274: 303-313.
LACHAUD, A., MAHE, G., 2008. Contribution à la connaissance de la diversité des abeilles sauvages de Loire-Atlantique. Bretagne vivante, SEPNB, 91 pp.
LECLERCQ, J., GASPAR, C., MARCHAL, J.-L., VERSTRAETEN, C., WONVILLE, C., 1980. Analyse des 1600 premières cartes de l'Atlas provisoire des Insectes de Belgique, et première liste rouge d'Insectes menacés dans la faune belge. Notes fauniques de Gembloux 4: 1-104.
LOUADI, K., 1999a. Systématique, éco-éthologie des abeilles (Hymenoptera, Apoidea) et leurs relations avec l'agrocénose dans la région de Constantine. Thèse Doc. Etat, sci.natu., Univ. Mentouri, Conctantine, 202 pp.
LOUADI, K., 1999b. Contribution à la connaissance des genres Halictus et Lasioglossum de la région de Constantine (Algérie) (Hymenoptera, Apoidea, Halictidae). Bull. Soc. Ent. France,104(2): 141–144.
LOUADI, K., DOUMANDJI, S.E., 1998a. Diversité et activité de butinage des abeilles (Hymenoptera, Apoidea) dans une pelouse à Thérophytes de Constantine (Algérie). Canadian Entomologist 130: 1–12.
LOUADI, K., DOUMANDJI, S.E., 1998 b. Note d'information sur l'activité des abeilles (domestiques et sauvages) et l'influence des facteurs climatiques sur les populations. Sci. Et Tech. Univ. Constantine 9: 83- 87.
LOUADI, K., BENACHOUR, K.,BERCHI, S., 2007a. Floral visitation patterns during spring in Constantine, Algeria. African Entomology 15(1): 209–213.
LOUADI, K., MAGHNI, N., BENACHOUR, K., BERCHI, S., AGUIB, S, MIHOUBI, I., 2007b. Présence de Dasypoda maura Pérez, 1895(Hym., Apoidea, Melittidae). Bull. Soc. Ent. Fance. 112(2): 232.
LOUADI, K., TERZO, M., BENACHOUR, K., BERCHI, S., AGUIB, S., MAGHNI, M. BENARFA, N., 2008. Les Hyménoptères Apoidea de l'Algérie orientale avec une liste d'espèces et comparaison avec les faunes ouest-paléarctiques. Bulletin de la Société entomologique de France 113 s (4): 459-472.
LOUVEAUX, J., 1980. Les abeilles et leur élevage. Ed. Hachette, Paris, 230 pp.
MAYER, C., KUHLMANN, M., 2004. Synchrony of pollinators and plants in the winter rainfall area of South Africa - observations from a drought year. Transactions of the Royal Society of South Africa 59: 55-57.
McGREGOR, S.E., 1976. Insect pollination of cultivated crop plants. Agriculture Handbook, Serv. Rech. Agri., U.S. Gov. Printing Off., Washington, (496): 411.
MICHENER, C.D., 1979. Biogeography of the bees. Annals of the Missouri Botanical Garden66: 77–347
MICHENER, C.D., 2007. The Bees of the World. second edition. Baltimore, 913 pp.
MICHEZ, D., TERZO, M., RASMONT, P ., 2004c. Phylogénie, biogéographie et choix floraux des abeilles oligolectiques du genre Dasypoda Latreille 1802 (Hymenoptera: Apoidea: Melittidae). Ann. Soc. Entomol. Fr., (n. s.)40(3-4): 421-435.
MONSEVIÈIUS, V., 2004. Comparison of three methods of sampling wild bees (Hymenoptera, Apoidea) in Èepkeliai Natural Reserve (South Lithuania). Ekologija 4: 32-39.
MÜLLER, A., KUHLMANN, M., 2008. Pollen hosts of western palearctic bees of the genus Colletes (Hymenoptera: Colletidae) – The Asteraceae paradox. Biological Journal of the Linnean Society (95): 719-733.
OERTLI, S., MÜLLER, A., DORN, S., 2005. Ecological and seasonal patterns in the diversity of a species-rich bee assemblage (Hymenoptera: Apoidea: Apiformes). Eur. J. Entomol. 102: 53-63.
PARKER, F.D., 1981. Sunflower pollination : abundance, diversity and seasonaly of bees and their effect on seed yelds. J. Apic. Res. 20(1): 49–61.
PATINY, S., MICHEZ, D., 2007. Biogeography of bees (Hymenoptera, Apoidea) in Sahara and the Arabian deserts. Insect Syst. Evol. 38: 19-34.
PEETERS, T.M.J., REEMER, M., 2001. Bijenfauna en beheer van zeven terreinen van Natuurmonumenten. Leiden: Stichting European Invertebrate Survey - Nederland, 67.
PESSON, P., LOUVEAUX, J., 1984. Pollinisation et production végétale. Ed. Institut national recherche agronomique, Paris, 637 pp.
PITKÄNEN, M., TIAINEN, J., 2001. Biodiversity of agricultural landscapes in Finland. BirdLife Conservation Series (No. 3). BirdLife; Finland.
POTTS, S.G., VULLIAMY, B., ROBERTS, S., O'TOOLE, C., DAFNI, A., NEEMEN, G., WILLMER, P., 2005. Role of nesting resources in organising diverse bee communities in a Mediterrranean landscape. Ecol. Entomol . 30: 78-85.
RAMADE, F., 1984. Eléments d'écologie. Ecologie fondamentale. Ed. Mc Graw-Hill, Paris, 397 pp.
RASMONT, P., MERSCH, P., 1988. Première estimation de la dérive faunique chez les Bourdons de la Belgique (Hymenoptera, Apidae). Annls Soc. r. Zool. Belg. 118(2): 141-147
RASMONT, P., BARBIER, Y., PAULY, A., 1990. Faunistique des Hyménoptères Apoïdes de deux terrils du Hainaut occidental. Notes fauniques de Gembloux 21: 39-58.
RASMONT, P., LECLERCQ, J., JACOB-REMACLE, A., PAULY, A., GASPAR, C., 1993. The faunistic drift of Apoidea in Belgium, p. 65-87. In : Bruneau, E. (Eds), Bees for pollination. Commission of the European Communities, Brussels, 237 pp.
RASMONT, P., EBMER, P.A., BANASZAK, J., VAN DER ZANDEN, G., 1995. Hymenoptera Apoidea Gallica. Liste taxonomique des abeilles de France, de Belgique, de Suisse et du Grand- duché de Luxembourg. Bull. Soc. Ent. France, 100 (hors série),pp. 1-98.
RASMONT, P., PAULY, A., TERZO, M., PATINY, S., MICHEZ, D., ISERBYT, S., BARBIER, Y., HAUBRUGE, E., 2005. The survey of wild bees (Hymenoptera, Apoidea) in Belgium and France. FAO, Roma, 18 pp.
ROTH, P., 1923. Contribution à la connaissance des Hyménoptères Aculeata de l'Afrique du Nord. Description de Bombex handirschella Ferton. Bulletin de la Société d'Histoire Naturelle de l'Afrique du Nord 14(5): 189–191.
ROTH, P., 1924. Contribution à la connaissance des Hyménoptères Aculeata de l'Afrique du Nord. 2. Note. Bulletin de la Société d'Histoire Naturelle de l'Afrique du Nord 15(3): 122–123.
ROTH, P., 1930. Hyménoptères recueillis au Sahara central par la mission scientifique du Hoggar. Bulletin de la Société d'Histoire Naturelle de l'Afrique du Nord 21(6–7): 79–86.
SAUNDERS, E., 1901. Hymenoptera Aculeata collected in Algeria. Part I – Heterogyna and Fossores to the end of Pompilidae, Trans. Ent. Soc. Lond 4: 515–525.
SAUNDERS, E., 1908. Hymenoptera Aculeata collected in Algeria. Part III – Anthophila. Transactions of the British Entomology Society 2: 177–273.
SCHULTHESS, A., 1924. Contribution à la con-naissance de la faune des Hyménoptères de l'Afrique du Nord. Bulletin de la Société d'Histoire Naturelle de l'Afrique du Nord 15(6): 93–320.
SELTZER, P., 1946. Le climat d'Algérie. Université d'Alger, Alger, 219 pp.
SONET, M., JACOB-REMACLE, A., 1987. Pollinisation de la légumineuse fourragère Hedysarum coronarium L. en Tunisie. Bull. Rech. Agro. Gembloux 22(1): 19–32.
SOUTHWOOD, T.R.E., 1978. Ecological methods. With particular reference to the study of insect populations. Ed. Chapman et Hall, London, 35 pp.
STALLEGGER, P., LIVORY, A., 2008. Inventaire et analyse du peuplement d'abeilles sauvages (Hymenoptera: Apidae) de l'Espace Naturel Sensible «Rives de Seine Sud», Berville-sur-Mer, Fatouville-Grestain, Fiquefleur-Equainville.Rapport de recherche, Conseil général de l'Eure - Direction du développement économique et de l'aménagement du territoire - Espaces naturels sensibles, Evreux (France), 65 p.
STEFFAN-DEWENTER, I., KLEIN, A.M., ALFERT, T., GAEBELE, V., TSCHARNTKE, T., 2006. Bee diversity and plant–pollinator interactions in fragmented landscapes, in: Waser N.M, Ollerton J. (Eds), Plant-pollinator Interactions, From specialization to generalization. The University of Chicago Press, Chicago, pp. 387–408.
STEWART, P., 1969. Quotint pluviométrique et degradation biosphérique. BulL. Doc. Hist. nat. agro. pp. 24–25.
TASEI, J.N., 1976. Les insectes pollinisateurs de la fèverole d'hiver (Vicia faba equina L.) et la pollinisation des plantes mâles – stériles en production de semences hybrides. Apidologie 7(1): 1–38.
WILLIAMS, P., 1982. The distribution and decline of British bumble bees (Bombus Latr.). Journal of Apicultural Research 21: 236-245.
Entregue para publicação em Julho de 2008
Aceite para publicação em Abril de 2010
