










Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiência e Técnica Vitivinícola
versão impressa ISSN 0254-0223
Ciência Téc. Vitiv. vol.27 no.2 Dois Portos dez. 2012
Densidade do coberto vegetal na casta Alfrocheiro (Vitis vinifera L.) I. Efeitos na estrutura e microclima do coberto vegetal, vigor e expressão vegetativa
Canopy density on the vine variety Alfrocheiro (Vitis vinifera L.) I. Effects on canopy structure, , microclimate, vigour and vegetative growth
Manuel Botelho*, Amândio Cruz, Rogério de Castro
Instituto Superior de Agronomia, Tapada da Ajuda, 1349-017 Lisboa, Portugal
*Corresponding author:
RESUMO
A avaliação dos efeitos da densidade de sarmentos e da desfolha sobre o microclima, estrutura do coberto vegetal, vigor e expressão vegetativa foi avaliada durante 3 anos (2004 a 2006), na casta Alfrocheiro, na região demarcada do Dão, Centro-Norte de Portugal. Ao estado fenológico G de Baggiolini (cachos separados) foram introduzidas três densidades de sarmentos: testemunha (D1), 18 sarmentos/m sebe (D2) e 12 sarmentos/m sebe (D3). Ao pintor foi introduzido o factor desfolha na zona dos cachos: F1 – com desfolha; F0 – sem desfolha. A nível fisiológico, em nenhum dos anos se encontraram diferenças significativas entre as diversas modalidades, no entanto, entre os três anos encontram-se diferenças apreciáveis na evolução sazonal do potencial hídrico foliar de base, maioritariamente explicadas pelas condições meteorológicas ocorridas, nomeadamente pela quantidade e distribuição da precipitação ao longo da estação de crescimento vegetativo. A resposta vegetativa das plantas foi amplamente marcada pelo ano, em especial pelo regime pluviométrico. Os crescimentos secundários (netas) em 2005, resultado do forte stress hídrico observado, foram reduzidos e como consequência a área foliar total das plantas foi inferior ao verificado em 2004 e 2006. Em qualquer dos anos, as diferenças encontradas na área foliar por cepa foram devidas ao números de sarmentos e não à área foliar por sarmento. O peso médio do sarmento aumentou proporcionalmente à redução do seu número, sem que tenha havido diferenças significativas no peso de lenha de poda por videira. A correção do número de sarmentos teve como consequência a redução da densidade do coberto vegetal na zona dos cachos, traduzida pelo menor número de camadas de folhas e pelo aumento da radiação intercetada. Efeito similar mas mais acentuado foi obtido pela desfolha. A nível vegetativo, embora pequenas variações tenham sido observadas entre as diversas densidades de sarmentos, em nenhuma delas se encontraram condições de ensombramento que tenham conduzido a situações de senescência foliar por falta de insolação.
Palavras-chave: densidade de sarmentos, desfolha, área foliar, estrutura do coberto, microclima e vigor.
SUMMARY
The evaluation of shoot density and basal leaf removal effects on canopy structure, canopy microclimate and on vigour and vegetative growth was carried out during 3 years (2004 to 2006) on Alfrocheiro vine variety, at Dão Delimited Region, Centre-North of Portugal. Three different shoot densities were assayed (at G stage of Baggiolini scale): control (D1), 18 shoots.m-1 row (D2) and 12 shoots.m-1 row (D3). At veraison, leaf removal was introduced at fruit zone – LR1 (no leaf removal - LR0). In any of the years were found significant differences between the different treatments, in terms of ecophysiological behaviour, however, between the three years significant differences in the seasonal evolution of predawn leaf water potential were observed, mostly explained by meteorological conditions, mainly the amount and distribution of rainfall during the growing season. The vegetative growth was largely marked by year, especially by rainfall. In 2005, the growth of lateral shoots, due to the strong water stress observed, was reduced and, as a consequence, the total leaf area per plant was lower than in 2004 and 2006. In any year, the differences in total leaf area per plant were due to the number of shoots, since the leaf area per shoot was equal. The shoot weight was increased proportionally to the reduction of its number, while no significant differences in pruning weight per vine were found.Shoot thinning resulted in a reduction of the canopy density at cluster zone, fewer leaf layer number and increased light interception. Similar but more pronounced effects were obtained by leaf removal. At the upper part of canopy, although minor variations were observed between different shoot densities, in none of them were observed shade conditions which have led to situations of leaf senescence by lack of sunlight.
Key words: shoot density, leaf removal, leaf area, canopy structure, microclimate and vigour.
INTRODUCÃO
As intervenções em verde são o conjunto de operações sobre os órgãos herbáceos da videira capazes de modificar o seu número, peso, superfície e posição (Branas, 1974). Estas intervenções são mais importantes quando o vigor das plantas é excessivo, levando à aglomeração de lançamentos e à distribuição não uniforme da área foliar e, consequentemente, ao ensombramento exagerado das uvas (Percival et al., 1994). Nestes casos, as técnicas de gestão do coberto vegetal, tornam-se uma ferramenta imprescindível para aumentar a qualidade das uvas (Smart et Robinson, 1991).
Uma das intervenções possíveis para manipular a densidade do coberto é a redução da densidade de sarmentos que, apesar de não ser correntemente usada, tem grande importância, pois influencia de forma directa a estrutura do coberto vegetal, modificando o seu microclima (Castro et al, 2007). As elevadas densidades de sarmentos permitem criar uma maior área foliar no início do ciclo (Miller et al., 1996), no entanto originam também mais sinks vegetativas para os hidratos de carbono, o que se traduz em mudanças significativas na estrutura do coberto, na redução do tamanho dos sarmentos, na redução do tamanho das folhas e da área foliar por sarmento Naor et al. (2002). Relativamente à área foliar, apesar de esta ser maior no início do ciclo, os estudos realizados não são coerentes quanto ao seu desenvolvimento durante o ciclo vegetativo, pois se Downton Grant (1992) referem uma recuperação, por parte das baixas densidades de sarmentos chegando mesmo a ultrapassar as altas densidades, Sommer Clingeleffer (1993) observaram uma manutenção dessas diferenças ao longo de todo o ciclo.
A redução da densidade do coberto permite um aumento da Radiação Fotossinteticamente Ativa (PAR) que penetra no interior do coberto, nomeadamente na zona dos cachos (Smart Robinson, 1991; Dokoozlian Kliewer, 1995). No entanto, dada a forte capacidade de adaptação das folhas ao ensombramento, que, segundo Heuvel et al. (2004), se pode refletir num abaixamento do ponto de compensação para a luz de 61%, e a sua boa adaptabilidade à radiação intermitente (Kriedemann, 1968), a assimilação fotossintética no interior do coberto pode assumir um papel de relevo, nomeadamente em regiões de clima mediterrânico, onde a elevada temperatura das folhas expostas (Kriedemann, 1968) e o stress hídrico (Flexas et al., 1999, Baeza et al., 2007) podem levar a reduções muito significativas da sua taxa fotossintética.
Foram já vários os estudos de optimização da densidade de sarmentos em várias castas. Basler (1980, 1981) concluiu que a densidade ideal para a Müller-Thurgau e a Blauburgunder, conduzidas em monoplano vertical ascendente, seriam cerca de 12 a 14 e 14 a 17 sarmentos/m de linha, respetivamente. Kiefer Crusius (1984) cit. in Reynolds et al. (1994) recomendam 21 a 29, 14 a 22 e 14 a 17sarmentos/m de linha para as castas Riesling, Müller-Thurgau e Silvaner, respetivamente. Com base em dados de microclima do coberto, Smart (1988) concluiu que 15 sarmentos/m de linha seria o ideal para a Gewürztraminer, ainda que estes dados tenham sido colhidos em vinhas com sebes muito densas, cuja correção da densidade foi feita ao pintor, pelo que o efeito do crescimento de netas não foi observado. Reynolds et al. (1994) concluíram que densidades entre 16 e 26 sarmento/m de linha são recomendáveis para a obtenção de quantidade e qualidade economicamente aceitáveis na casta Riesling.
A desfolha é outra das intervenções em verde passível de ser usada na redução da densidade do coberto e consiste na remoção de um número variável de folhas na zona de frutificação, aumentando a exposição dos cachos à luz solar e o seu arejamento, prevenindo doenças criptogâmicas e facilitando a vindima (Smart Robinson, 1991). A desfolha ao pintor não traz perdas a nível da fotossíntese já que remove as folhas mais velhas e portanto com menor capacidade fotossintética (Vasconcelos e Castagnoli, 2000; Cruz et al., 2001). Por outro lado, quando realizada mais precocemente, observa-se um aumento da taxa fotossintética das folhas remanescentes (Candolfi-Vasconcelos et al., 1994) que pode compensar a área foliar perdida.
A casta utilizada neste estudo, Alfrocheiro, muito disseminada na região do Dão, está na base de alguns dos melhores vinhos desta região e atualmente começa a ser plantada, com bons resultados, noutras regiões portuguesas, como é o caso do Alentejo. É uma casta de porte ereto, com uma boa capacidade produtiva e de maturação precoce (Lopes et al., 2008; Eiras-Dias et al., 2011) tendo, no entanto, uma grande sensibilidade a ataques de Botrytis cinerea Pers., os quais em alguns anos podem provocar grandes perdas de rendimento e qualidade. Assim, com este trabalho pretende-se estudar o efeito de intervenções na densidade do coberto e otimizar a densidade dos sarmentos para a casta Alfrocheiro, cultivada nas condições edafoclimáticas da região do Dão, contribuindo desta forma para aumentar o arejamento dos cachos, reduzir a intensidade dos ataques de Botrytis cinerea Pers. e otimizar a produtividade e a qualidade das uvas e dos vinhos.
MATERIAL E MÉTODOS
Caracterização da parcela experimental e do clima
A vinha, onde foi realizado o ensaio, localiza-se em Carregal do Sal, na Região Demarcada do Dão (Dão Sul, Soc. Vitivinícola, S.A.). A casta Alfrocheiro foi enxertada em 1991, sobre 1103 P, conduzida em Cordão Royat bilateral e a vegetação em monoplano vertical ascendente. O compasso é de 2,5x1,2m, estando as linhas orientadas no sentido N-S.
De acordo com o balanço hídrico de Thornthwaite, o clima desta região é mesotérmico, com concentração da eficiência térmica na estação quente, nula ou pequena, sub-húmido a seco com um moderado excesso de água no Inverno (B3 a C1s). Os dados climáticos foram recolhidos na estação meteorológica do Centro de Estudos Vitivinícolas de Nelas. O solo é, de acordo com a classificação da FAO-UNESCO, um Cambissolo, franco-arenoso, de origem granítica, com reacção ácida e baixa reserva hídrica.
O delineamento experimental é do tipo split-split-plot com 3 repetições. Foram ensaiadas 3 densidades de sarmentos (introduzidas no estado G da escala de Baggiolini): Testemunha (D1), 18 sarmentos.m-1 (D2) e 12 sarmentos.m-1 (D3) e ao pintor foi introduzida a desfolha: F1 – com desfolha; F0 – sem desfolha e a monda de cachos: M1 - com monda (remoção de aproximadamente 35% dos cachos); M0 - sem monda. Em 2004, não foi introduzida a monda de cachos, pelo que o delineamento foi do tipo split-plot com 3 repetições.
Na modalidade D1, pretendeu-se simular o desladroamento mecânico, ou seja, foram retirados apenas os ladrões que se desenvolveram sobre o tronco da planta. Neste caso, permaneceram na planta os ramos ladrões originados nos braços e os ramos normais provenientes das unidades de frutificação. Em D2, foram retirados todos os ramos ladrões do tronco e braços da planta, de modo a que o seu número fosse de 18 sarmentos por metro linear de sebe. Finalmente em D3, para além do desladroamento do tronco e braços, sobre as unidades de produção foram retirados abrolhamentos múltiplos e pâmpanos menos desenvolvidos de modo a que por metro linear de sebe ficassem 12 sarmentos.
Ao pintor, foi introduzida a desfolha (F1) que consistiu na remoção das folhas basais até à folha oposta ao cacho mais basal.
Em 2005 e 2006, a monda de cachos foi feita com base na sua qualidade potencial, retirando todos os cachos mais atrasados e/ou de sarmentos com menor vigor. No entanto, como verificámos que a monda não influenciou o microclima, a estrutura do coberto vegetal, nem o vigor, esses resultados não são aqui apresentados.
Estado hídrico e trocas gasosas
O potencial hídrico foliar de base (ψb) foi determinado com uma câmara de pressão, como descrito por Scholander et al. (1965), ao longo do ciclo vegetativo, até próximo da vindima. Em cada densidade de sarmentos foram colhidas, em videiras diferentes, 8 folhas adultas, bem expostas e do terço médio dos sarmentos.
A medição das trocas gasosas ao nível dos estomas foi realizada com um sistema portátil (modelo ADC-LCA4). Estas medições foram efectuadas ao longo do ciclo vegetativo em 6 folhas em cada modalidade de densidade de sarmentos, situadas no terço médio do coberto vegetal, bem expostas e em condições atmosféricas estáveis (céu limpo).
Como a desfolha não influenciou o ψb nem as trocas gasosas, os seus resultados não serão apresentados.
Estrutura e microclima do coberto vegetal
A estrutura e o microclima do coberto vegetal foram caracterizados, nos 3 anos, durante a maturação.
Para a determinação da área foliar usou-se a metodologia proposta por Lopes e Pinto (2005). Em cada modalidade foram eleitos 6 sarmentos normais, frutíferos e de vigor médio.
A estimativa da Superfície Externa do Coberto Vegetal (SECV), aqui designada por Superfície Foliar Exposta (SFE), proposta por Murisier e Zufferey (1997) foi obtida a partir do perímetro exposto, tendo sido realizadas 60 medições em cada modalidade com recurso a uma régua graduada de 2,5m de comprimento.
A densidade do coberto vegetal foi avaliada através do Número de Camadas de Folhas (NCF), seguindo o método Point Quadrat proposto por Smart e Robinson (1991). Em cada modalidade, fizeram-se 60 inserções perpendiculares à parede de vegetação, com uma fina vareta metálica, ao nível dos cachos e ao nível vegetativo (0,6m acima da zona de frutificação). Com os dados obtidos foi calculado o NCF e a percentagem de folhas e de cachos interiores.
A quantificação da Radiação Fotossinteticamente Ativa (PAR) intercetada pelo coberto, foi realizada com recurso a um ceptómetro (SunScan Ceptometer-type SS1 - Delta-T Devices) para a medição da PAR no interior do coberto vegetal, ao nível dos cachos e a nível vegetativo. O ceptómetro foi inserido, em cada nível, longitudinalmente no centro do coberto vegetal e paralelamente ao solo. Foram realizadas medições a meio da manhã (10h), zénite solar e a meio da tarde (17h), de forma a avaliar a evolução da interceção de radiação ao longo do dia. A cada hora foram feitas 24 medições em cada modalidade e em cada nível do coberto.
Vigor
Durante a poda, para avaliar o vigor e a expressão vegetativa dos diferentes tratamentos, contabilizaram-se o número de varas existentes em cada videira e o seu peso. Em cada modalidade foi contabilizada a lenha de poda de 12 videiras, previamente eleitas e equilibradas.
Como a desfolha não influenciou os parâmetros do vigor, os seus resultados não são apresentados.
Análise Estatística
A análise dos resultados relacionados com comportamento ecofisiológico (potencial hídrico foliar de base, fotossíntese e taxa de transpiração) foi corrigida pelo erro padrão da média com recurso ao programa MOExcel. Os restantes resultados foram sujeitos a uma análise de variância com recurso ao programa Statistica 6.0.
RESULTADOS E DISCUSSÃO
Clima
Analisando os dados climáticos dos 3 anos em estudo (Figura 1), podemos constatar que em 2004, nos meses de Julho, Agosto e Setembro, a temperatura média mensal foi inferior à média de 30 anos, contrariamente a Junho que foi um mês mais quente que a média. Em 2005, as temperaturas em Junho e Agosto foram superiores à média, enquanto que nos restantes meses do ciclo vegetativo, foram idênticas. Em 2006, a temperatura foi superior à média de 30 anos desde o início do ciclo e, globalmente, este foi o ano mais quente dos três.
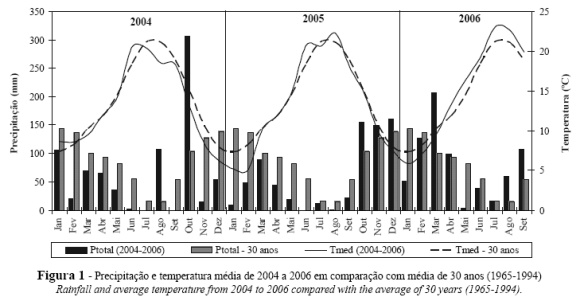
Em termos de precipitação, 2004 foi um ano menos chuvoso que a média, sendo a única excepção Agosto em que ocorreu uma pluviosidade muito superior à média (cerca de 107 mm vs 15 mm).
No ano de 2005, os valores de precipitação foram inferiores à média não só no Inverno, mas também durante todo o ciclo vegetativo da vinha, condicionando significativamente a reserva hídrica do solo.
Em 2006, a precipitação total, ocorrida entre Janeiro e Setembro, foi superior à ocorrida durante o mesmo período de 2005 (705 vs 243 mm) e próxima da média da região (705 vs 695 mm) o que levou a uma boa reposição das reservas hídricas do solo o Outono-Inverno (847 mm entre Out. 2005 e Mar. 2006). Durante o ciclo vegetativo da vinha, só nos meses de Maio e Junho foram observados valores de precipitação inferiores à média, enquanto que nos meses de Agosto e Setembro foram superiores.
Estado hídrico da vinha
O estado hídrico da vinha ao longo do período vegetativo foi avaliado através do potencial hídrico foliar de base (ψb) e é apresentado na Figura 2. Esta variável, reflete o potencial de água do solo na zona explorada pelas raízes e existem atualmente valores de referência consistentes que correlacionam ao valores obtidos com os níveis de stress a que as plantas estão submetidas em cada fase do ciclo e/ou ano (Carbonneau, 2001; Ojeda, 2001; Deloire et al., 2003).
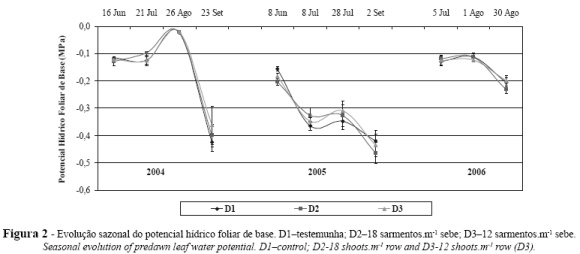
Em qualquer dos anos é possível observar que não existem diferenças significativas ao nível do potencial hídrico foliar de base entre as densidades de sarmentos. Assim, embora se verifiquem grandes diferenças entre a área foliar total das videiras (vide Tabela I) e mesmo na área foliar mais reativa e portanto com maior taxa transpiratória (Carbonneau, 1995), a superfície foliar exposta (SFE – Tabela II), estas não se traduziram no esgotamento da reserva hídrica na zona radicular. Resultados idênticos foram observados por Andrade (2003) e Cruz et al. (2012).
Entre os três anos encontram-se diferenças apreciáveis na evolução sazonal do ψb, maioritariamente explicadas pelas condições meteorológicas verificadas, nomeadamente pela quantidade e distribuição da precipitação ao longo da estação de crescimento vegetativo. De qualquer modo durante a floração (final do mês de Maio), em qualquer dos anos, as plantas encontravam-se sem restrições hídricas, visto os valores de Yb serem superiores a -0,2 MPa (Riou e Payan, 2001; Deloire et al., 2003).
Em 2004, a precipitação ocorrida durante o Inverno e nos meses de Abril e Maio permitiram o excelente abastecimento das plantas em água até Julho. À recuperação observada a 26 de Agosto, devida à ocorrência de chuva entre 1 e 19 desse mês (106,6 mm), seguiu-se um decréscimo acentuado até à vindima (24 de Setembro) para valores próximos de -0,4 MPa, situação de défice hídrico ligeiro a moderado, considerada como favorável à obtenção de uvas de qualidade (Carbonneau, 2001; Ojeda, 2001; Deloire et al., 2003). Este decréscimo foi devido à ausência de precipitação desde 19 de Agosto até essa data, associada a temperaturas elevadas.
Nos dois anos seguintes, é de assinalar que, devido às condições meteorológicas durante o Inverno e a Primavera serem tão distintas (entre 1 de Janeiro e 30 de Junho a precipitação acumulada em 2005 foi de 208mm e em 2006 de 523mm), a reserva hídrica do solo no início do Verão (8 de Julho de 2005 e 5 de Julho de 2006) foi totalmente díspar. Assim, neste período, em 2005 verificou-se uma situação de stress hídrico médio, enquanto que em 2006, os valores observados correspondem a uma situação de stress hídrico nulo (Deloire et al., 2003). Durante o Verão a diferença entre os dois anos acentuou-se, devido à pouca precipitação ocorrida em 2005, durante Julho e Agosto (14mm), comparativamente com igual período de 2006 (75mm).
Em 2005 as reservas hídricas do solo, na zona radicular, foram decrescendo gradualmente até à data da vindima, tendo-se aqui registado valores considerados óptimos, por Ojeda (2001) e Deloire et al. (2003), para a produção de uvas tintas para vinhos de qualidade, concentrados, equilibrados e aptos para envelhecimento. Em 2006, o ψb sofreu um ligeiro decréscimo durante a fase de maturação, no entanto, este foi o ano com maior disponibilidade hídrica do solo, situando-se este parâmetro em valores considerados de stress hídrico nulo.
Trocas gasosas
A acumulação de matéria seca numa vinha está amplamente dependente da fotossíntese do coberto vegetal, que é fortemente determinada pela área foliar e pela sua capacidade para interceptar e usar a radiação solar (Dokoozlian e Kliewer, 1995).
A evolução sazonal da taxa fotossintética e da taxa transpiratória ao longo do período vegetativo nos três anos pode ser observada na Figura 3. De um modo geral, em qualquer dos anos, constata-se que as diferenças entre densidades de sarmentos são variáveis e, normalmente, sem significado estatístico, não havendo, portanto, uma correlação aparente entre a densidade do coberto vegetal e o comportamento ecofisiológico das plantas. Este resultado está, aliás, em consonância com a ausência de diferenças verificada no potencial hídrico foliar de base. Valores similares foram encontrados por Cavallo et al. (2001), ao avaliarem também as trocas gasosas em folhas expostas da casta Aglianico conduzida de três formas distintas.
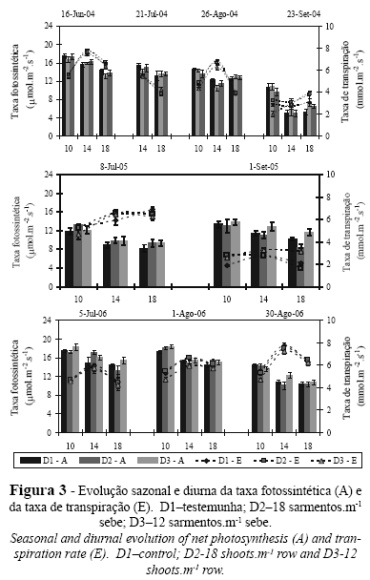
As diferenças nas trocas gasosas foram principalmente sentidas entre anos e muito marcadas pelo regime hídrico das plantas. Relativamente a 2004, os valores das taxas transpiratória e fotossintética mantêm-se em valores elevados até 26 de Agosto, refletindo a ausência de restrições hídricas que se verificou até esta data (Figura 2). De 26 de Agosto para 23 de Setembro observa-se uma queda acentuada nestas taxas, intimamente correlacionada com o decréscimo da disponibilidade hídrica na zona radicular. Vários trabalhos realizados em condições mediterrânicas referem a estreita correlação entre a disponibilidade hídrica na rizosfera e as trocas gasosas ao nível dos estomas (Lopes, 1994; Lopes, 1998; Souza et al. 2003; Hera-Orts et al., 2004). O gradual envelhecimento das folhas conduz também à perda da capacidade fotossintética, pois estamos perante folhas com mais de 50 dias de idade (Poni et al., 1994).
Também em 2005, se observa a redução acentuada da taxa de transpiração de 8 de Julho para 1 de Setembro, reflexo da menor disponibilidade hídrica do solo, visto o ψb ter atingido nesta data o valor mais negativo dos 3 anos (ca. -0,45 MPa). No entanto, a taxa fotossintética manteve-se a níveis elevados (ca. 10-12 µmol.m-2.s-1). Em 2006, a chuva ocorrida ao longo do ciclo vegetativo, manteve as plantas em conforto hídrico (Figura 2) o que resultou em valores elevados da taxa de transpiração, enquanto que a fotossíntese sofreu um decréscimo até à vindima devido ao envelhecimento da folha (Poni et al., 1994).
Em qualquer dos anos, a casta Alfrocheiro manteve taxas fotossintéticas relativamente elevadas, em condições de disponibilidade hídrica diferentes. Desta forma, parece-nos que esta casta tem uma grande capacidade de adaptação a diferentes condições ambientais, nomeadamente ao clima mediterrâneo e às previsíveis alterações climáticas.
Estrutura do coberto vegetal
No Quadro I são apresentados os dados relativos à área foliar nos três anos do ensaio. Analisando a área foliar principal por sarmento, constata-se que esta aumentou com a diminuição da densidade de sarmentos em 2004 e 2005. No entanto, em 2006 não houve diferenças significativas entre densidades. Os resultados obtidos em 2004 e 2005, anos mais secos, estão em consonância com os obtidos por Naor et al. (2002), Intrigliolo e Castel (2011) e Myers et al. (2008), enquanto que em 2006 são similares aos obtidos por Reynolds et al. (1994). O aumento da área foliar principal por sarmento deveu-se principalmente ao aumento da área das folhas, já que o número de folhas permaneceu inalterado (dados não apresentados), visto a desponta ter sido feita à mesma altura para todas as modalidades.
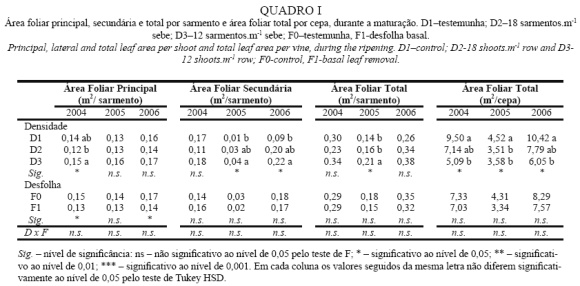
A desfolha conduziu a uma redução da área foliar principal por sarmento em 2004 e 2006, a qual se deveu à redução do número de folhas principais por sarmento. Em 2005, tal como observado por Howell et al. (1994), apesar de ter havido uma tendência para a redução da área foliar principal por sarmento, provocada pela desfolha, os resultados não foram estatisticamente significativos.
Relativamente à área foliar secundária, observa-se o seu maior desenvolvimento nas densidades mais baixas, em 2005 e 2006, como verificado também por Smart e Robinson (1991) e Reynolds et al. (1994). Por outro lado, verifica-se que em 2004 e 2006 houve um maior desenvolvimento de netas que em 2005, que resulta da maior disponibilidade hídrica existente nestes dois anos o que originou um aumento do vigor (ver Quadro VIII). O facto de em 2006 ter havido um aumento da diferença entre D1 e as restantes densidades, prende-se, provavelmente, com as condições climáticas deste ano (maior disponibilidade hídrica e térmica - ver Figura 1), que conduziram a uma emissão de ladrões superior a 2004 e 2005, reduzindo o vigor individual de cada sarmento (ver Quadro VIII). Resultados idênticos foram observados por Chaves et al. (2007).
Ainda no Quadro I pode observar-se que nos três anos em estudo houve uma tendência para um acréscimo da área foliar por sarmento com a diminuição da densidade de sarmentos e naturalmente o aumento individual do seu vigor (ver Quadro VIII), ainda que só em 2005 existam diferenças estatísticas. Resultados idênticos foram observados por Intrigliolo e Castel (2011), Miller et al. (1996). A desfolha não provocou diferenças significativas da área foliar por sarmento em nenhum dos anos, contrariamente ao encontrado por vários autores (Andrade, 2003; Correia, 2012). Este facto deve-se à existência de folhas senescentes na modalidade F0, as quais não foram contabilizadas na medição da área foliar mas, no entanto, influenciam o NCF e o microclima luminoso na zona dos cachos.
Finalmente, analisando a área foliar por cepa, verifica-se que apesar da menor área foliar por sarmento, a densidade D1 apresentou maiores valores de área foliar por planta, em qualquer dos anos, devido ao maior número de sarmentos. Tendo em conta os valores observados, verifica-se que estes corroboram os obtidos por Naor et al. (2002) e contrariam os de Sommer e Clingeleffer (1993) e Myers et al. (2008), já que, apesar de a correção da densidade de sarmentos ter sido feita antes da floração, o que, de acordo com Reynolds et al. (2005), permite à planta recuperar melhor a área folia r perdida, até ao final do ciclo, D1 apresentou sempre uma área foliar por cepa superior às restantes densidades. Em 2005, para além de D2 e D3 terem uma área foliar por cepa muito semelhante entre si, D1 não foi muito superior em termos de valor absoluto, sugerindo que num ano em que o regime hídrico leva a uma redução do vigor, este torna-se o principal factor limitante na produção de área foliar, pois nestes anos o aumento da densidade de sarmentos, reduz significativamente a área foliar por sarmento.
Relativamente à influência da desfolha sobre a área foliar, não houve uma redução significativa da área foliar por cepa em nenhum dos anos.
Finalmente, mais do que a influência das modalidades de gestão do coberto, o factor preponderante na área foliar das videiras foi o ano. Efectivamente, em 2005, ano em que as reservas hídricas do solo e as taxas de produção de fotoassimilados das plantas durante a fase de crescimento vegetativo foram menores, houve um menor desenvolvimento de área foliar das videiras, em qualquer das modalidades. A diminuição fez-se sentir mais ao nível da área foliar secundária, sintoma de um menor vigor de acordo com Smart e Robinson (1991), a qual levou a uma menor área foliar total por cepa.
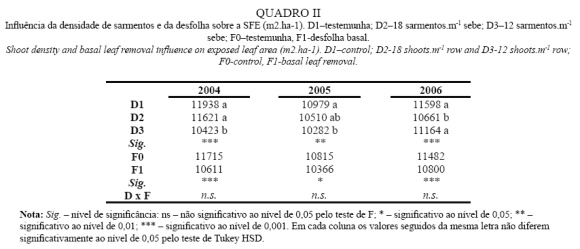
Superfície Foliar Exposta
Analisando a SFE, apresentada no Quadro II, podemos verificar que entre os três anos não houve diferenças assinaláveis, o que se deve ao facto do método utilizado na avaliação desta variável, levar em conta apenas as dimensões da sebe, as quais não se alteraram significativamente, já que foram controladas mecanicamente. No entanto, observam-se diferenças significativas entre modalidades em todos os anos. Os valores obtidos em todas as situações são, de acordo com Argillier (1989), indicativos de uma vinha de qualidade, quando conduzida em monoplano ascendente e em clima mediterrânico.
Ao nível da densidade de sarmentos, verifica-se que as diferenças entre densidades não são coerentes nos três anos do ensaio. Em 2004 e 2005, observa-se uma redução da SFE com a redução da densidade de sarmentos. Já em 2006, constata-se uma redução significativa da SFE em D2, sendo D1 e D3 idênticas.
Ao nível da desfolha, observa-se um decréscimo significativo da SFE com a realização da desfolha, facto que tem na sua origem a redução da altura da sebe, devido à remoção das folhas basais. No entanto, dado as folhas removidas serem as mais velhas e portanto com uma menor capacidade fotossintética (Poni et al., 1994), a redução na produção global de fotoassimilados pelas plantas não tem repercussão na evolução da maturação (Kriedeman et al., 1970; Castro et al., 2006).
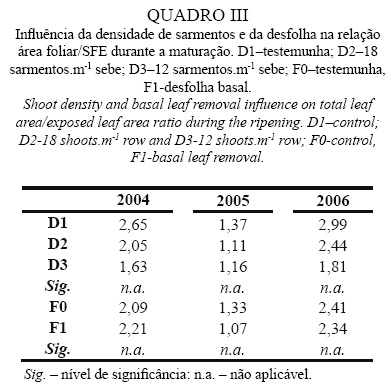
Relação Área Foliar / Superfície Foliar Exposta
No Quadro III pode observar-se que a influência da densidade de sarmentos e da desfolha, na relação entre a área foliar total (AF) e a SFE não foi igual nos três anos. Globalmente, os valores desta relação foram superiores em 2004 e 2006, o que é devido à maior expansão vegetativa observada nestes anos, a qual levou a uma maior densidade do coberto e, consequentemente, a uma maior proporção de folhas ensombradas.
Por outro lado, constata-se que a redução da densidade de sarmentos, foi mais eficiente a diminuir a relação AF/SFE nos anos de maior desenvolvimento vegetativo (2004 e 2006), ainda que em 2005 se observe uma ligeira tendência neste sentido. Ao reduzir a densidade de sarmentos, diminui-se a proporção de folhas interiores, referida como potenciadora da eficiência fotossintética por Carbonneau (1995) e Wolf et al. (1986), já que a SFE tem, em condições normais, maior capacidade fotossintética que a superfície foliar ensombrada.
A desfolha não conduziu a alterações relevantes deste parâmetro em nenhum dos anos em estudo, provavelmente por terem sido removidas folhas ensombradas e expostas em proporções idênticas.
De um modo geral, qualquer das modalidades em estudo apresenta-se mais equilibrada nesta relação (AF/SFE) em 2005, já que se encontram abaixo de 1.5, situação considerada ideal por Smart e Robinson (1991).
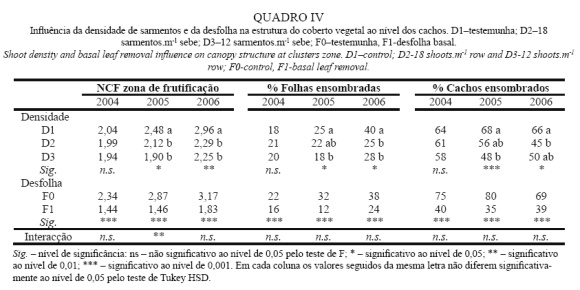
Densidade do Coberto Vegetal
A densidade do coberto é apresentada no Quadro IV, onde se pode observar que a correção da densidade de sarmentos originou uma redução significativa da densidade do coberto em 2005 e 2006, patente na diminuição do NCF, à qual correspondeu um decréscimo da percentagem de folhas e cachos ensombrados. Estes dados vão de encontro aos observados por Reynolds et al. (2005). No entanto, de acordo com os dados apresentados no Quadro IV, verifica-se que, em 2005, a redução do NCF aconteceu somente nas modalidades desfolhadas, não tendo a correção da densidade de sarmentos, por si só, originado a redução deste parâmetro. Em todos os anos, incluindo 2004, a desfolha originou um efeito mais eficiente na redução da densidade do coberto e na melhoria do microclima dos cachos, do que a correção da densidade de sarmentos, tendo originado diferenças significativas em todos os parâmetros apresentados. Também Percival et al. (1994) e Andrade (2003) obtiveram reduções significativas da densidade do coberto com a aplicação de desfolha.
Na origem da maior efetividade da desfolha na redução da densidade do coberto, está, provavelmente, o facto de a correção da densidade de sarmentos ser realizada precocemente e de serem removidas sinks ativas, levando portanto a mudanças importantes na distribuição de fotoassimilados e permitindo uma maior taxa de crescent dos sarmentos remanescentes. Desta forma, tal como referido por Smart e Robinson (1991), houve um maior desenvolvimento das netas nas menores densidades de sarmentos (conforme Quadro I) que impediram reduções da densidade do coberto mais significativas. No caso da desfolha, dado que foi realizada mais tarde, numa fase de baixo ou nulo crescimento vegetativo, e tendo sido removidas folhas com menor capacidade fotossintética (Vasconcelos e Castagnoli, 2000; Cruz et al., 2001), não houve alterações na distribuição de fotoassimilados. Consequentemente, nesta fase do ciclo vegetativo a planta não respondeu de forma a compensar a área foliar removida pela desfolha tendo, portanto, a densidade do coberto sido efetivamente reduzida.
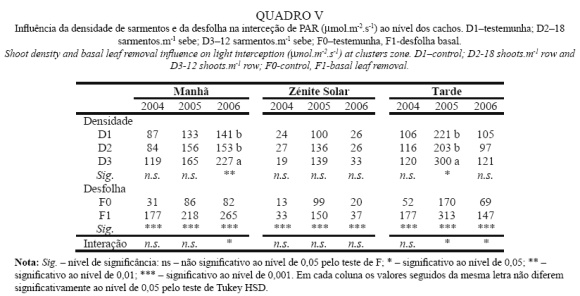
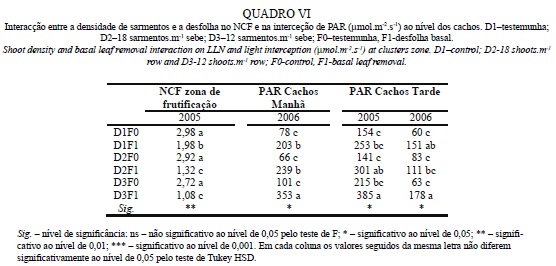
O Quadro V apresenta os dados relativos à PAR intercetada na zona dos cachos em 3 fases do dia: Manhã, Zénite Solar e Tarde. Relativamente à influência da densidade de sarmentos, nota-se que não é estatisticamente significativa em nenhuma das medições, pois apesar de ter havido diferenças nas medições da manhã em 2006 e na da tarde em 2005, no Quadro VI, constata-se que entre as modalidades não desfolhadas não houve diferença significativas entre densidades, tendo sido a desfolha a originar uma maior PAR ao nível dos cachos em D3.
Analisando o efeito da desfolha, verifica-se que aumentou drasticamente a PAR na zona dos cachos em todas as medições e em todos os anos. Este resultado decorre da redução do NCF apresentada no Quadro IV e vai de encontro ao obtido por Bledsoe et al. (1988), Payan (1997) e Andrade et al. (2001).
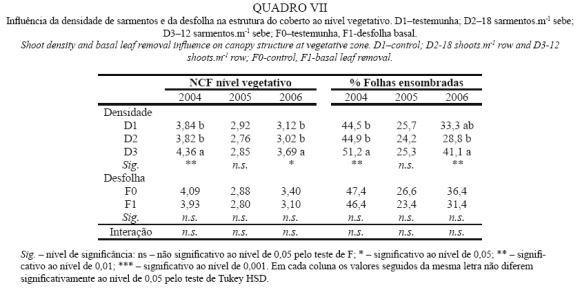
A densidade do coberto ao nível vegetativo é apresentada no Quadro VII, verificando-se tendência para o aumento do NCF a nível vegetativo com a redução da densidade de sarmentos. A causa deste aumento está na resposta da planta à redução do número de sarmentos com a emissão de netas, a qual está patente na área foliar secundária apresentada no Quadro I. Assim, os anos em que este efeito é significativo são os de maior vigor e em que as diferenças entre a área foliar secundária são menores.
Paralelamente, a percentagem de folhas ensombradas e, portanto, com menor taxa fotossintética aumenta com a redução da densidade de sarmentos. No entanto, os níveis de PAR no interior do coberto são superiores ao ponto de compensação para a luz da fotossíntese (Chaves, 1986) em qualquer dos anos (dados não apresentados), o que mostra que apesar da mais baixa taxa fotossintética destas folhas é superior a zero, não prejudicando a produção global de fotoassimilados da planta.
A desfolha não teve nenhuma influência no NCF ao nível vegetativo, tal como também não teve na área foliar secundária em qualquer dos anos.
Vigor e Expressão Vegetativa
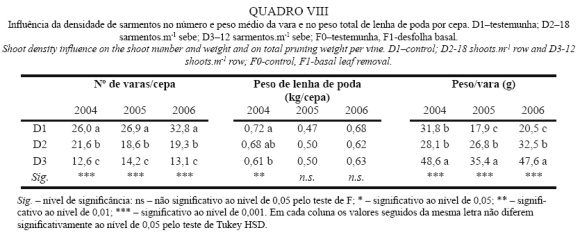
No Quadro VIII apresenta-se a influência da correção da densidade de sarmentos no vigor e na expressão vegetativa. Através da sua análise, observa-se em primeiro lugar que a correção da densidade de sarmentos, realizada numa fase precoce do ciclo (cachos separados), foi eficiente no controlo da densidade de sarmentos. De facto, as modalidades D2 e D3 apresentaram valores, deste parâmetro, idênticos nos três anos e próximos do que se pretendia.
Em qualquer dos anos na maior densidade de sarmentos não se verificaram problemas de atempamento das varas, contrariamente ao observado por Reynolds et al. (1994) na casta Riesling, com densidades idênticas às implementadas neste estudo.
Analisando o peso de lenha de poda por cepa, pode verificar-se que não existiram diferenças significativas entre densidades de sarmentos, excepto em 2004.
Relativamente ao peso médio da vara, observa-se o efeito depressivo do constrangimento hídrico que se verificou em 2005 sobre este parâmetro, que limitou a atividade fisiológica das plantas, tendo sido desenvolvida uma menor área foliar tanto a nível do sarmento como da cepa (Quadro I). Analisando os dados das diferentes modalidades em estudo, constata-se o aumento do vigor individual do sarmento, com o decréscimo do seu número. Tendo em conta a manutenção da produção global de lenha de poda, este resultado era espectável e está de acordo com o observado por Miller et al. (1996) e por Reynolds et al. (1994).
Analisando os valores obtidos em qualquer dos anos à luz dos considerados por Smart e Robinson (1991), como indicativos de uma vinha em equilíbrio (20 a 40g/vara), verifica-se antes de mais que, com excepção de D3 em 2004 e 2006 e D1 em 2005, todas a modalidades se encontraram dentro do referido intervalo, evidenciando uma situação de equilíbrio da vinha. Relativamente, às densidades extremas, observa-se uma aproximação aos limites do intervalo. Já a densidade intermédia, é aquela que revela uma situação de maior equilíbrio, tendo-se situado o peso da vara sempre na zona central do intervalo.
CONCLUSÕES
As intervenções em verde em estudo, de um modo geral, não influenciaram de forma significativa o comportamento fisiológico das plantas. Contudo, entre os três anos encontram-se diferenças apreciáveis na evolução sazonal do potencial hídrico foliar de base e da taxa fotossintética, essencialmente explicadas pelas condições meteorológicas verificadas, nomeadamente pela quantidade e distribuição da precipitação ao longo da estação de crescimento vegetativo.
Em termos de estrutura do coberto vegetal observou-se também um efeito marcado do ano, em especial o regime hídrico. Os crescimentos secundários (netas) em 2005, resultado do forte stress hídrico observado, foram muito reduzidos e como consequência a área foliar total das plantas foi muito inferior ao verificado em 2004 e 2006. Em qualquer dos anos, as diferenças encontradas na área foliar por cepa foram devidas, essencialmente, ao números de sarmentos e não à área foliar por sarmento. Assim, as densidades de sarmentos mais baixas apesar de menor área foliar por cepa, apresentaram maior proporção de folhas expostas.
A supressão de sarmentos teve como consequência a redução da densidade do coberto vegetal na zona dos cachos, traduzida pelo menor número de camadas de folhas e pelo aumento da radiação intercetada. A nível vegetativo, embora pequenas variações tenham sido observadas entre as diversas densidades de sarmentos, em nenhuma delas se encontraram condições de ensombramento que tenham conduzido a situações de senescência foliar por falta de insolação.
Apesar de não ter havido diferenças significativas na maior parte dos anos no peso de lenha de poda por videira, o peso médio do sarmento aumentou proporcionalmente à redução do seu número, tendo D2 apresentado os resultados mais equilibrados, ao longo dos 3 anos.
A desfolha não influenciou de forma significativa a área foliar por cepa. No entanto, reduziu de forma muito efetiva a densidade do coberto, a qual se refletiu num menor NCF e num aumento muito significativo da PAR intercetada ao nível dos cachos.
Concluindo, a correção da densidade de sarmentos, por ter sido feita numa fase de crescimento ativo da vegetação, permitiu o reequilíbrio da videira, esbatendo os efeitos na redução da densidade do coberto e melhoria do microclima dos cachos. Por outro lado, a desfolha, por ter sido realizada ao pintor, teve maior efeito na alteração da estrutura do coberto vegetal.
AGRADECIMENTOS
À Agência de Inovação pelo financiamento do Projecto LUSOCASTAS, ao abrigo do qual foi realizado este estudo. Ao CEVDão pela cedência dos dados climáticos. A todos os estagiários que colaboraram na colheita de dados.
REFERÊNCIAS BIBLIOGRÁFICAS
Andrade I., Pedroso V., Martins S., Brites J., Lopes C., 2001. Efeito de diferentes intensidades de desfolha na produção e na qualidade do mosto da casta Jaen. 5º Simpósio de Vitivinicultura do Alentejo, Évora, Portugal, vol.2: 1 – 7. [ Links ]
Andrade I., 2003. Efeito da intensidade da desfolha da videira (Vitis vinífera L.) na fotossíntese, na produção e na qualidade. 215p. Dissertação de Doutoramento. Instituto Superior de Agronomia, Universidade Técnica de Lisboa. [ Links ]
Argillier J. P., 1989. Interdependance des facteurs de la qualité. Chambre dAgriculture de lHérault, Montpellier. [ Links ]
Baeza P., Sanchez-de-Miguel P., Centeno A., Junquera P., Linares R., Lissarrague J. R., 2007. Water relations between leaf water potential, photosynthesis and agronomic vine response as a tool for establishing thresholds in irrigation scheduling. Scientia Horticulturae, 114, 151-158. [ Links ]
Basler P., 1980. Optimal pruning of RieZur optimalen Belastung der Reben beim Riesling x Sylvaner am Zurichsee. Schweiz Z. Obst-und Weinbau, 116, 56-64. [ Links ]
Basler P., 1981. Zur optimalen Belastung der Reben beim Blauburgunder in der Ostschweiz. Schweiz Z. Obst-und Weinbau, 117, 354-364. [ Links ]
Bledsoe A.M., Kliewer W.M., Marois J.J., 1988. Effects of timing and severity of leaf removal on yield and composition of Sauvignon Blanc grapevines. Am. J. Enol. Vitic., 39, 49-55. [ Links ]
Branas J., 1974. Viticulture. 990 p. Ed. Dehan, Montpellier. [ Links ]
Candolfi-Vasconcelos M.C., Koblet W., Howell G.S., Zweifel W., 1994. Influence of Defoliation, Rootstock, Training System, and Leaf Position on Gas Exchange of Pinot Noir Grapevines. Am. J. Enol. Vitic. 45, 173-180. [ Links ]
Carbonneau A., 2001. Gestion de leau dans le vignoble: Théorie et pratique. 3-21. In: 12èmes Journées GESCO. Montpellier, France. [ Links ]
Carbonneau A., 1995. La surface foliaire exposée potentielle. Guide pour sa mesure.39-48. 8èmes Journées GESCO, Vairão, Portugal. [ Links ]
Castro R., Cruz, A., Botelho M., 2006. Tecnologia Vitícola. 160p. Ministério da Agricultura, Pescas e Florestas/Direcção Geral de Agricultura da Beira Litoral/Comissão Vitivinícola da Bairrada, Coimbra. [ Links ]
Castro R., Cruz A., Rodrigues C.A., Botelho M., Rodrigues C., Rodrigues A., Gomes C., 2007. Shoot density and basal leaf removal effects on microclimate, yield, fruit composition and wine quality of the portuguese vine variety Touriga Nacional. 798-811. In: 15èmes Journées GESCO, Porec, Croatia. [ Links ]
Cavallo P., Poni S., Rotundo A., 2001. Ecophysiology and vine performance of cv. Aglianico under various training systems. Scientia Horticulturae, 87, 21-32. [ Links ]
Chaves M., 1986. Fotossíntese e repartição dos produtos de assimilação em Vitis vinífera L. 220 p. Dissertação de doutoramento, Instituto Superior de Agronomia, Universidade Técnica de Lisboa. [ Links ]
Chaves M.M., Santos T.P., Souza C.R., Ortuño M.F., Rodrigues M.L., Lopes C.M., Maroco J.P., Pereira J.S., 2007. Deficit irrigation in grapevine improves water-use efficiency while controlling vigour and production quality. Ann. Appl. Biol., 150, 237-252. [ Links ]
Correia J., 2012. Efeito da época e da intensidade de desfolha na casta Sauvignon. 91p. Dissertação de Mestrado. Instituto Superior de Agronomia, Universidade Técnica de Lisboa. [ Links ]
Cruz A., Almeida C., Frade P., Ribeiro F., Aires A., Castro R., 2001. Controlo da produção e do vigor pela carga e intervenção em verde cv. Arinto, Bairrada. Vol. 1, 217-224. In Actas do 5º Simpósio de Vitivinicultura do Alentejo. [ Links ]
Cruz A., Botelho M., Silvestre J., Castro R., 2012. Soil management: introduction of tillage in a vineyard with a long-term natural cover. Ciência Téc. Vitiv., 27, 27-38. [ Links ]
Deloire A, Carbonneau A., Federspiel B., Ojeda H., Wang Z., Costanze P. 2003. La vigne et leau. Progrés Agricole et Viticole, 120, 79-90. [ Links ]
Dokoozlian NK, Kliewer WM., 1995. The light environment within grapevine canopies. I. Description and seasonal changes during fruit development. Am. J. Enol. Vitic., 46, 209–218. [ Links ]
Downton W.J.S., Grant W.J.R., 1992. Photosynthetic physiology of spur pruned and minimal pruned grapevines. Austral. J. Plant Physiol, 19, 309-316. [ Links ]
Eiras-Dias J., Faustino R., Clímaco P., Fernandes P., Cruz A., Cunha J., Veloso M., Castro R., 2011. Catálogo das castas para vinho cultivadas em Portugal. Vol. 1, 109 p. Instituto da Vinha e do Vinho. [ Links ]
Flexas J., Escalona J. M., Medrano H., 1999. Water stress induces different levels of photosynthesis and electron transport rate regulation in grapevines. Plant, Cell and Environment, 22, 39-48. [ Links ]
Hera-Orts M.L., Martínez-Cutillas A., López-Roca J. M., Gómez-Plaza E., 2004. Effects of moderate irrigation on vegetative growth and productive parameters of Monastrell vines grown in semiarid conditions. Span. J. Agric. Res., 2, 273-281. [ Links ]
Heuvel J.E.V., Proctor J.T.A., Fisher K.H., Sullivan J.A., 2004. Shading affects morphology, dry-matter partitioning, and photosynthetic response of greenhouse-grown Chardonnay grapevines. HortScience, 39, 60-70. [ Links ]
Howell G.S., Candolfi-Vasconcelos M.C., Koblet W., 1994. Response of Pinot Noir grapevine growth, yield, and fruit composition to defoliation the previous growing season. Am. J. Enol. Vitic., 45, 188-191. [ Links ]
Intrigliolo D. S., Castel J. R. 2011. Interactive effects of deficit irrigation and shoot and cluster thinning on grapevine cv. Tempranillo. Water relations, vine performance and berry and wine composition. Irrig Sci., 29, 443-454. [ Links ]
Kiefer W., Crusius P., 1984. Beziehungungen zwischen Anschnitt, Mengenertrag und Qualität bei verschiedenen Rebsorten. Mitt. Klosterneuburg, 34, 51-63. [ Links ]
Kriedemann P.E., 1968. Photosynthesis in vine leaves as a function of light intensity, temperature, and leaf age. Vitis, 7, 213-220. [ Links ]
Kriedemann P.E., Kliewer W.M., Harris J.M., 1970. Leaf age and photosynthesis in Vitis vinifera L. Vitis, 9: 97-104. [ Links ]
Lopes, C., 1998. Relationships between leaf water potential and photosynthetic activity of field-grown grapevines under a mediterranean environment. Acta Hort., 493, 287-292. [ Links ]
Lopes C. M. A., 1994. Influência do sistema de condução no microclima do coberto, vigor e produtividade da videira (Vitis vinifera L.). 205p. Tese de Doutoramento. Universidade Técnica de Lisboa. Instituto Superior de Agronomia. [ Links ]
Lopes C.M.A., Pinto P.A., 2005. Easy and accurate estimation of grapevine leaf area with simple mathematical models. Vitis, 44, 55-61. [ Links ]
Lopes J., Eiras-Dias J.E., Abreu F., Clímaco P., Cunha J.P., Silvestre J., 2008. Exigências térmicas, duração e precocidade de estados fenológicos de castas da colecção ampelográfica nacional. Ciência Téc. Vitiv., 23, 61-71. [ Links ]
Miller, D.P., Howell, G.S., Flore, J.A., 1996. Effect of Shoot Number on Potted Grapevines: I. Canopy Development and Morphology. Am. J. Enol. Vitic., 47, 244-250. [ Links ]
Murisier F., Zufferey V., 1997. Rapport feuille-fruit de la vigne et qualité du raisin. Revue Suisse Vitic. Hortic., 29, 355-362. [ Links ]
Myers J.K., Wolpert J.A., Howell G.S., 2008. Effect of shoot number on the leaf area and crop weight relationship of young Sangiovese grapevines. Am. J. Enol. Vitic., 59 (4): 422-424. [ Links ]
Naor A., Gal Y., Bravdo B., 2002. Shoot and cluster thinning influence vegetative growth, fruit yiel, and wine quality of Sauvignon blanc grapevines. J. Amer. Soc. Hort. Science, 127, 628-634. [ Links ]
Ojeda H., 2001. Bases ecophysiologiques et choix tecniques dans la gestion de leau dans les vignobles dArgentine. In: Actes 12ème Journées GESCO-Journée professionelle "Gestion d`eauans le vignoble", 75-86. [ Links ]
Payan J., 1997. Les travaux en vert: incidence sur la conduite de la vigne et sur la qualité de la récolte. In: EUROVITI 97. 11e Colloque et nologique Viticulure-nologie. [ Links ]
Percival D.C., Fisher K.H., Sullivan J.A., 1994. Use of fruit zone leaf removal with Vitis vinifera L. cv. Riesling grapevines. I. Effects on canopy structure, microclimate, bud survival, shoot density, and vine vigor. Am. J. Enol. Vitic., 45, 123-132. [ Links ]
Poni S., Intrieri C., Silvestroni O., 1994. Interactions of leaf age, fruiting, and exogenous cytokinins in Sangiovese grapevines under non-irrigated conditions. I. Gas exchange. Am. J. Enol. Vitic., 45, 71-78. [ Links ]
Reynolds A.G., Edwards C.G., Wardle, D.A., Webster D.R., Dever M., 1994. Shoot density affects Riesling grapevines I. Vine performance. J. Amer. Soc. Hort. Science, 119, 874-880. [ Links ]
Reynolds A.G., Molek T., Savigny C., 2005. Timing of shoot thinning in Vitis vinifera: Impacts on yield and fruit composition variables. Am. J. Enol. Vitic., 56, 343-356. [ Links ]
Riou C., Payan J.C., 2001. Outils de gestion de l'eau en vignoble méditerranéen. Application du bilan hydrique au diagnostique du stress hydrique de la vigne. In: Actes 12ème Journées GESCO- Journée professionelle "Gestion d`eau dans le vignoble",, 125-133. [ Links ]
Scholander P. F., Hammel H.T., Bradstreet E.T., Hemmingsen E.A., 1965. Sap pressure in vascular plants. Science, 148, 339-346. [ Links ]
Smart R.E., 1988. Shoot spacing and canopy light microclimate. Am. J. Enol. Vitic., 39, 325-333. [ Links ]
Smart R.E., Robinson M., 1991. Sunlight into wine. A Handbook for winegrape canopy management. Winetitles, Adelaide. [ Links ]
Sommer K. J., Clingeleffer P. R., 1993. Comparison of leaf area development, leaf physiology, berry maturation, juice quality, and fruit yield of minimal and cane pruned Cabernet sauvignon grapevines. In: Proceedings of the Second N. J. Shaulis Grape Symposium. [ Links ]
Souza C., Maroco J., Santos T., Rodrigues M., Lopes C., Pereira J., Chaves M., 2003. Partial rootzone drying: regulation of stomatal apperture and carbon assimilation in field-grown grapevines (Vitis vinifera cv. Moscatel). Functional Plant Biology, 30, 653-662. [ Links ]
Vasconcelos M.C., Castagnoli S., 2000. Leaf canopy structure and vine performance. Am. J. Enol. Vitic., 51, 390-396. [ Links ]
Wolf T.K., Pool R.M., Mattick L.R., 1986. Responses of young Chardonnay grapevines to shoot topping, ethephon on basal leaf removal. Am. J. Enol. Vitic., 37, 263-268. [ Links ]
*Corresponding author:
Tel.; +351 213 653 100, e-mail: mbotelho@isa.utl.pt
(Manuscrito recebido em 04.12.2012. Aceite para publicação em 14.01.2013)
